Серосодержащей аминокислотой является: серосодержащие аминокислоты | это… Что такое серосодержащие аминокислоты?
Что это — серосодержащие аминокислоты
Наверное, ни для кого не секрет, что аминокарбоновые кислоты (аминокислоты) – это бусины в сложной цепочке белков любого живого организма. Но мало кто знает, что практически все они были открыты во второй половине прошлого столетия.
Интересующие нас в этой статье биогенные серосодержащие аминокислоты – это цистеин и метионин. Попробуем обогатить знания читателя об этих двух органических соединениях с точки зрения биохимии и биологии нашего здоровья.
Кирпичики, из которых строится белок
Аминокислоты (серосодержащие в том числе) — довольно широко представленные в природе органические соединения. Сегодня нам известно более 500 аминокислот. При этом 240 из них встречаются в окружающей среде в свободном виде, а все остальные являются промежуточными продуктами обменных процессов в живых организмах.
И сегодня остается загадкой, почему из всего этого многообразия в белках человека встречается только 20 аминокислот (на фото выше). Они носят название биогенных, или участвующих в биосинтезе белка. Среди этих «избранных» только две являются серосодержащими аминокислотами. Это метионин (Methionine) и цистеин (Cysteine), в составе которых имеется атом серы.
Они носят название биогенных, или участвующих в биосинтезе белка. Среди этих «избранных» только две являются серосодержащими аминокислотами. Это метионин (Methionine) и цистеин (Cysteine), в составе которых имеется атом серы.
Отзывы: Метионин. Инструкция по применению, состав и…
«Метионин» является лекарственным средством с гепатопротекторным действием. Препарат содержит…
Все аминокислоты для нашего организма делятся на две группы: незаменимые (те, которые не синтезируются в организме человека) и заменимые (те, что организм может произвести сам). Цистеин относится ко второй группе, а вот метионин — к первой, которую мы должны получать с пищей — именно с ним связаны патологии, которые обусловлены нарушением обмена серосодержащих аминокислот. Но обо всем по порядку.
Что такое C5h21NO2S?
Метионин в чистом виде – это бесцветные кристаллы, довольно неприятно пахнущие и растворимые в воде. Это аминокислота, которая в нашем организме служит поставщиком метильных групп и серы.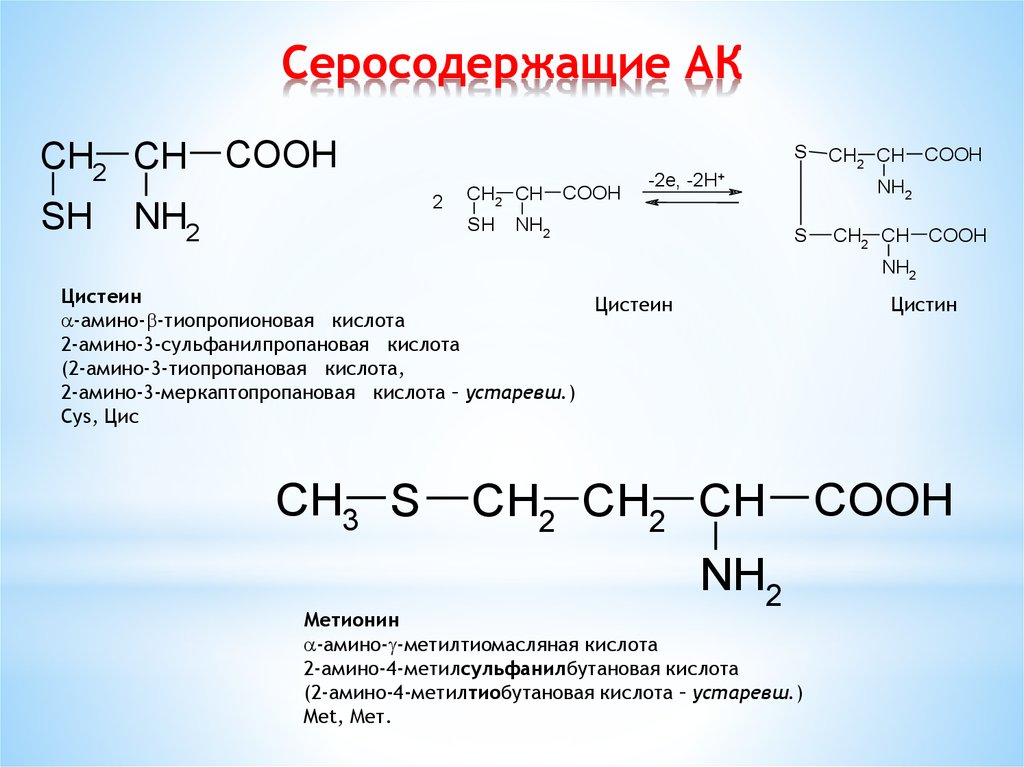 Как уже говорилось, метионин является незаменимой серосодержащей аминокислотой и не синтезируется в организме.
Как уже говорилось, метионин является незаменимой серосодержащей аминокислотой и не синтезируется в организме.
Он поступает с пищей и используется нашими клетками для собирания на рибосомах собственных белков. Эта серосодержащая аминокислота содержится в белке молока (казеин), в яйцах, кунжуте, муке, свинине и курятине, рыбе (лосось и тунец), в пшенице и овсе, в бобовых, во всех зеленых овощах, в сыре пармезан и моцарелла.
Кроме того, современная фармакология имеет в своем арсенале препараты, которые абсолютно идентичны метионину. Благодаря его роли в синтезе белка и жирорастворимым свойствам данные средства широко используются в спортивной сфере.
Отзывы: Метионин. Инструкция по применению, состав и…
«Метионин» является лекарственным средством с гепатопротекторным действием. Препарат содержит…
Значение метионина для организма
Биохимия обмена серосодержащих аминокислот поставляет нашему организму серу и метильные группы. Данная аминокислота играет значительную роль в синтезе цистеина, таурина, адреналина и мелатонина.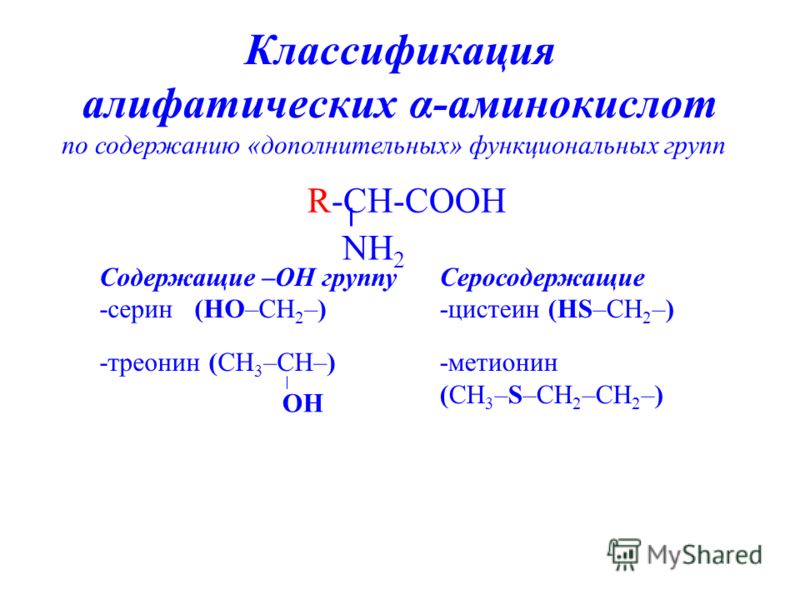
Функции ее в организме разнообразны и довольно широки:
- Участвует в снижении уровня холестерина и способствует профилактике атеросклероза.
- Выступает гепатопротектором – защищает печень от отложения излишков жира, восстанавливает ее структуру.
- Благотворно влияет на состояние почек и выделительной системы.
- Выступает в роли антидепрессанта, восстанавливает и нормализует сон, устраняет хроническую усталость.
- Улучшает состояние кожи, ногтей и волос.
- Способен обезвреживать токсические металлы, свободные радикалы и таким образом обеспечивать интоксикацию организма.
- Улучшает состояние суставов. Доказано, что у больных артритом, количество серы в хрящах уменьшено в 3 раза по сравнению с нормой.
И это далеко не полный перечень. Считается, что для обеспечения нашего организма этой аминокислотой, необходимо в сутки получать 19 мг метионина на 1 килограмм веса.
C3H7NO2S – протектор и очиститель
Цистеин – аминокислота, которая синтезируется в нашем организме из метионина. Процесс этот многоступенчатый и происходит при наличии специфических ферментов, витаминов. Сбой в нем может произойти на каждом этапе, и это также связано с патологиями обмена серосодержащих кислот.
Процесс этот многоступенчатый и происходит при наличии специфических ферментов, витаминов. Сбой в нем может произойти на каждом этапе, и это также связано с патологиями обмена серосодержащих кислот.
Аминокислоты — для чего нужны? Аминокислоты в продуктах….
Аминокислоты — одни из самых важных органических соединений. В этой статье мы расскажем, какими они…
Производными распада цистеина являются таурин и глутатион. Первое – белок, который отвечает за адекватную работу головного мозга, а второе играет важную роль в работе иммунной системы. Сам же цистеин влияет на состояние кожи (отвечает за синтез коллагена) и процессы детоксикации. Он входит также в состав многих ферментов пищеварительного тракта.
Значение для организма
Следует отметить такие положительные функции этой аминокислоты:
- Цистеин улучшает рост волос, обеспечивая синтез коллагена, что положительно влияет и на формирование кожи, структуру ногтей.
- Эта серосодержащая аминокислота влияет на формирование мышц и сжигание жира.

- Выполняет функции антиоксиданта, выводя токсины, особенно при одновременном приеме витамина С и селена.
- Активирует лейкоциты и лимфоциты, влияя на их противовирусные, противоопухолевые, противовоспалительные свойства.

Суточная норма цистеина для здорового взрослого человека – до 3 грамм в сутки. При этом повышение дозы может привести к токсическому отравлению.
Красивая цветная реакция
Определить наличие метионина и цистеина легко, и этот опыт часто показывают в старших классах школы. Реакция на серосодержащие аминокислоты (реакция Фоля) — одно из немногих именных, красивых и зрелищных химических преобразований.
Делается это так. В пробирку, где находится ацетат свинца (1 мл), добавляют раствор гидроксида натрия 10% и куриный белок. Затем смесь нагревают. В пробирке появляется черный осадок, и чем он темнее, тем больше серосодержащих аминокислот имеется в растворе.
В основе реакции лежит расщепление щелочью серы с образованием сероводорода, который связывается с натрием и выпадает в осадок.
Реакция широко применяется в клинико-биологических исследованиях, фармацевтике.
Дефицитные состояния
Гомоцистинурия – наследственная патология обмена серосодержащих аминокислот. Поражает нервную, кожно-мышечную, сосудистую системы, сопровождается отставанием в умственном развитии, судорогами, патологиями хрусталика глаза и зрительных нервов, нарушениями в формировании скелета и мышц.
Частота патологии — 1 случай на 100 тысяч новорожденных. В случае ранней диагностики данного вида энзимопатии лечение включает диету с исключением метионина и витаминную терапию.
Это не единственная патология обмена серосодержащих аминокислот, но остальные являются еще более редкими.
Приобретенные нарушения
Дефицитные состояния обычно проявляют себя в виде ломкости ногтей, выпадении волос, отечности мягких тканей, обострений болезней сердца, печени и почек.
Людям с повышенной кислотностью желудка прием продуктов, богатых метионином, практически противопоказан. Избыток его в организме вызывает аллергии, сонливость и расстройства пищеварительной системы.
Избыток его в организме вызывает аллергии, сонливость и расстройства пищеварительной системы.
А вот при стрессах и физических нагрузках наш организм требует увеличения серосодержащих аминокислот в рационе. Кроме того, беременным женщинам не стоит пренебрегать продуктами, богатыми ими. Это залог нормального формирования нервной системы у плода.
Помощник при похудении
В пятерке аминокислот, которые помогут сбросить лишний вес, цистеин занимает не последнее место.
Эта аминокислота сигнализирует мозгу о том, что желудок уже полон. Благодаря цистеину мы чувствуем себя сытыми. А это ведет к недопущению переедания, снижению веса и увеличению скорости похудения.
Ешьте брокколи, овсянку с бананами, яйца, морскую рыбу — и эта анорексигенная аминокислота поможет добиться стройности фигуры.
Метионин. Свойства, особенности, сфера применения
Метионин
CAS номер: 59-51-8
Брутто формула: C5h21NO2S
Внешний вид: порошок белого цвета со специфическим запахом
Химическое название и синонимы: DL-Methionine, DL-2-Amino-4-(methylthio)butyric acid; Acimetion
Физико-химические свойства:
Молекулярная масса: 149. 21 г/моль
21 г/моль
Плотность 1,34
Температура плавления 270-273 ° С
Температура кипения 186 ° C
рН 5,0-6,5 при 140 г / л при 25 ° C (77 ° F)
Растворимость в воде 2,9 г / 100 мл (20 ºC)
Описание:
Метионин является незаменимой аминокислотой, содержащей в своем составе атом серы. Является прекурсором цистеина (образуется из серина с участием метионина как серосодержащая аминокислота) и таурина (образуется из цистеина). Одно из наиболее важных превращений метионина — преобразование в S-аденозилметионин или «SAM». SAM участвует во многих разнообразных химических реакциях, перенося часть себя на другие молекулы, включая ДНК и белки. SAM также используется в производстве креатина — важной молекулы для клеточной энергии. Так как метионин в ряде биохимических реакций метаболизируется до большого спектра различных веществ, то тем самым прямо или косвенно он участвует во многих важных процессах, протекающих в организме.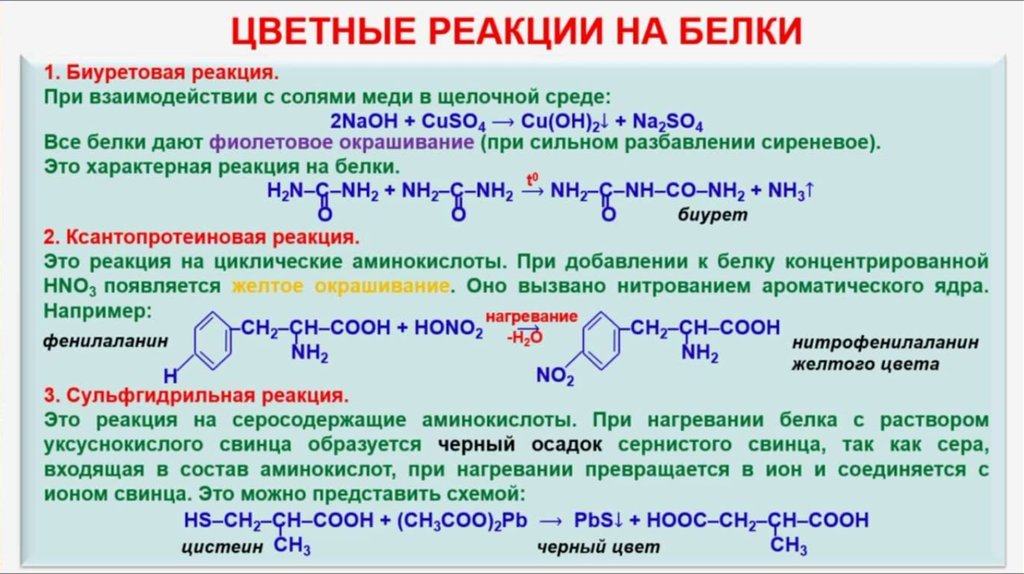 Кроме того, метионин играет важную роль в синтезе других белков, таких как карнитин или мелатонин.
Кроме того, метионин играет важную роль в синтезе других белков, таких как карнитин или мелатонин.
Метионин в крови позволяет сократить уровень гистамина, что в свою очередь уменьшает аллергические реакции организма. Обладает антиоксидантными и детоксицирующими свойствами, связываясь со свободными радикалами и токсинами. Метионин также обладает жирорастворимым эффектом и уменьшает осаждение жира в печени. Свойство метионина подкислять мочу позволяет использовать его при болезнях мочевыводящих путей и при образовании камней в почках.
Метионин является важным хрящеобразующим веществом. Для хряща в суставах требуется сера. Если в организме недостаточно серы, это может иметь негативные последствия для здорового человека в долгосрочной перспективе. Например, у людей, страдающих артритом, процесс заживления поврежденной ткани может быть длительным, если в начале болезни наблюдался дефицит серы.
Метионин не синтезируется организмом и должен поступать внутрь с пищей.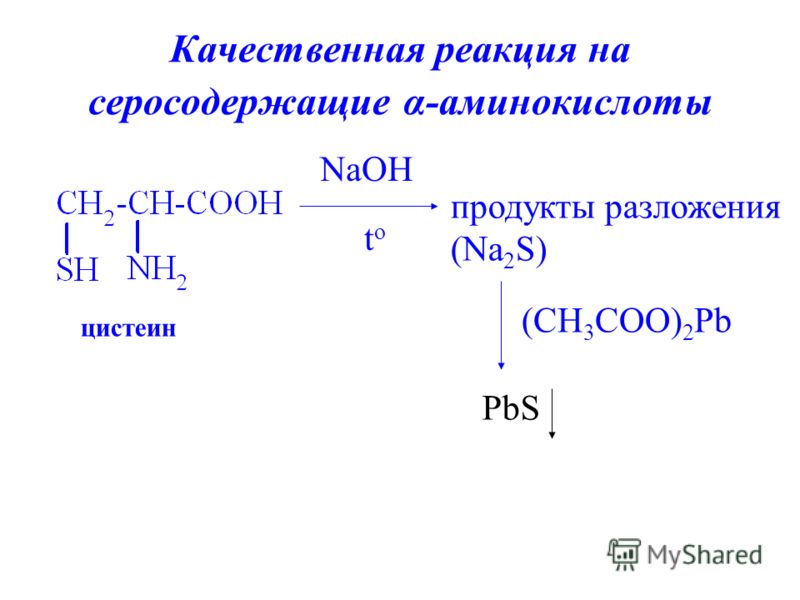 Источниками метионина служат мясо животных (говядина, курица и др), молочные продукты (йогурт,сыр и др), орехи (высокое содержание в бразильском орехе), яйца, бобовые, семена кунжута, тыквы, подсолнечника и др.
Источниками метионина служат мясо животных (говядина, курица и др), молочные продукты (йогурт,сыр и др), орехи (высокое содержание в бразильском орехе), яйца, бобовые, семена кунжута, тыквы, подсолнечника и др.
Применение:
Перорально метионин используется для предотвращения повреждения печени при отравлении ацетаминофеном и для тестирования людей на гипергомоцистеинемию. Он также применяется перорально для понижения pH мочи, лечения заболеваний печени, вирусных инфекций, включая вирус папилломы человека, вирус простого герпеса и опоясывающий лишай, снижения риска колоректального рака и / или рака молочной железы, уменьшения боли, вторичной по отношению к панкреатиту. Метионин используют перорально при врожденных дефектах нервной трубки, депрессии, алкоголизме, аллергии, астме, токсичности меди, побочных эффектах радиации, шизофрении, отмене лекарств, постменопаузальных состояниях, включая приливы, и болезни Паркинсона.
Метионин является БАДом для обогащения пищи людей, одним из важных компонентов в спортивном питании.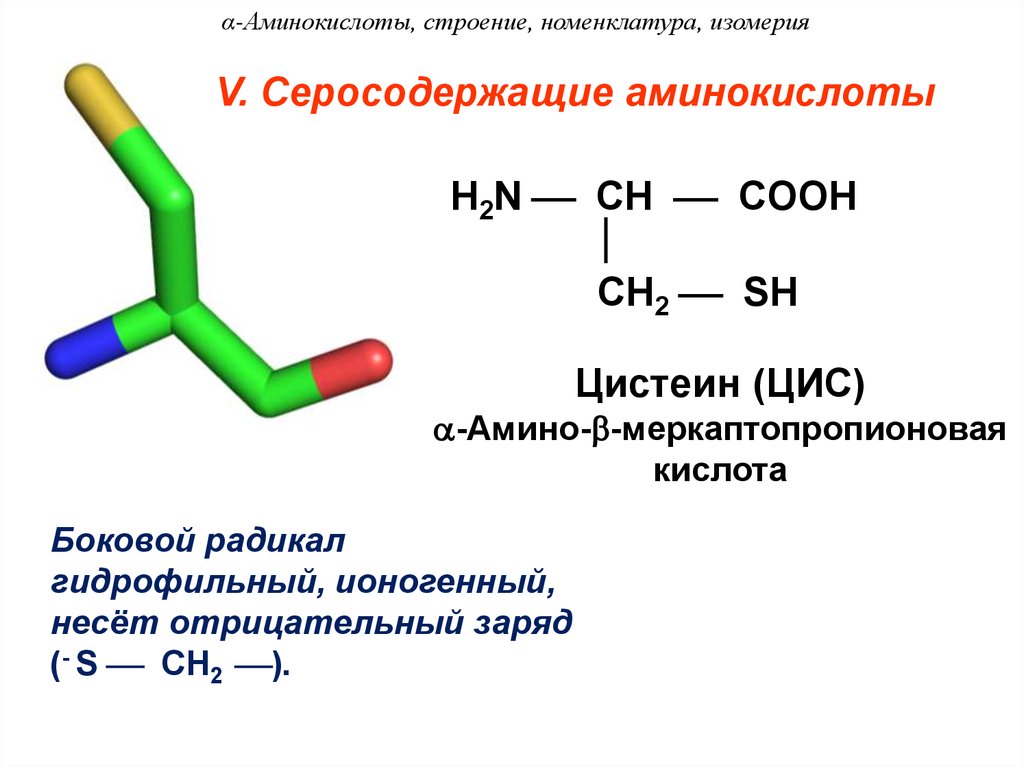
Получение:
Чистый способ получения D, L-метионина, включает следующие стадии: приготовление раствора цианида калия с использованием кристаллизованного маточного раствора, содержащего карбонат калия в качестве поглощающей синильную кислоту жидкости, затем взаимодействие раствора цианида калия с 3 -метилтиопропиональдегидом и раствора бикарбоната аммония при 50-150 ° С в течение 3-15 минут для получения раствора 5- (β-метилтиоэтил) гликолюрей, доведение раствора 5- (β-метилтиоэтил) гликолюрей до температуры 140-220 ° С и получение реакции омыления в течение 2-5 минут, после завершения омыления снижение температуры до 0-40 ° С, экстракция органическим растворителем, нейтрализация водной фазы с помощью СО 2 , кристаллизация, затем фильтрация, промывка и сушка с получением приемлемого продукта D, L-метионина; доведение кристаллизованного маточного раствора D, L-метионина от фильтрации до температуры 110-160 ° С для удаления СО2, который затем циркулируют и используют в качестве жидкости, поглощающей синильную кислоту.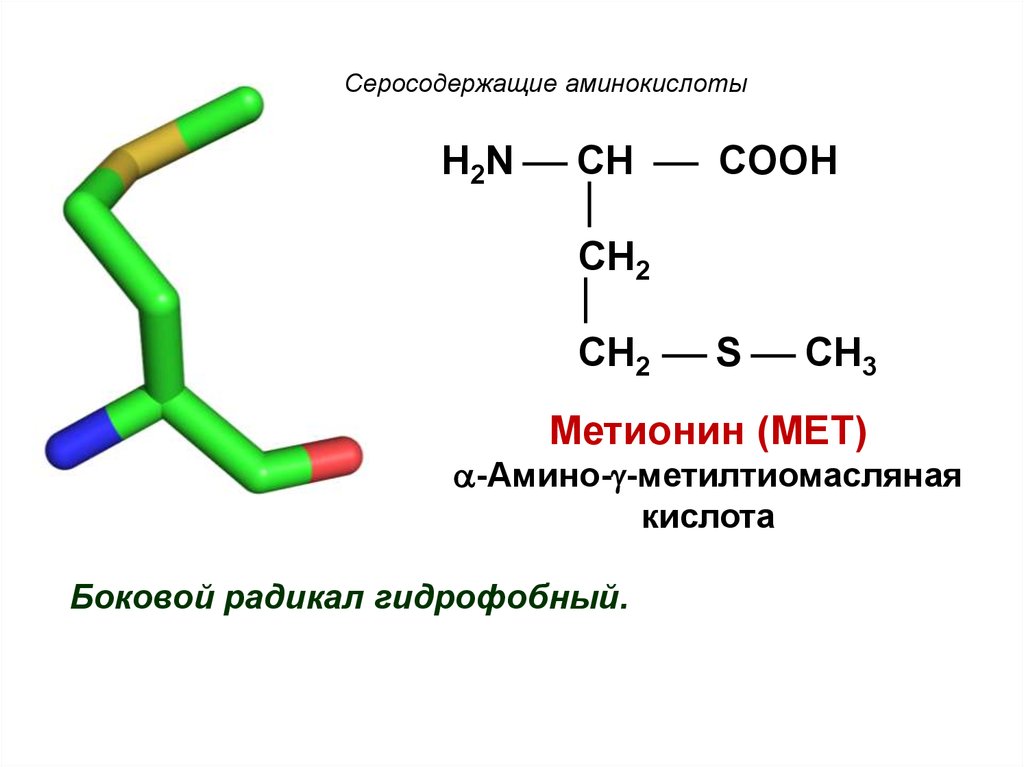 Технологический процесс настоящего изобретения представляет собой путь, подходящий для непрерывного и чистого производства, по существу, без производства сточных вод и отработанного газа.
Технологический процесс настоящего изобретения представляет собой путь, подходящий для непрерывного и чистого производства, по существу, без производства сточных вод и отработанного газа.
Используется метионин и в ветеринарии. Его иногда применяют как ингредиент корма для собак. Метионин может помочь снизить шансы возникновения камней у собак. Он разрешен в качестве дополнения к органическому корму для сельскохозяйственной птицы.
Метионин можно использовать как нетоксичный вариант пестицида против гигантских гусениц-ласточек, которые являются серьезным вредителем апельсиновых культур.
Действие на организм:
Механизм возможной антигепатотоксической активности L-метионина не совсем ясен. Считается, что метаболизм высоких доз ацетаминофена в печени приводит к снижению уровня глутатиона и усилению окислительного стресса. L-метионин является предшественником L-цистеина. Сам L-цистеин может обладать антиоксидантной активностью.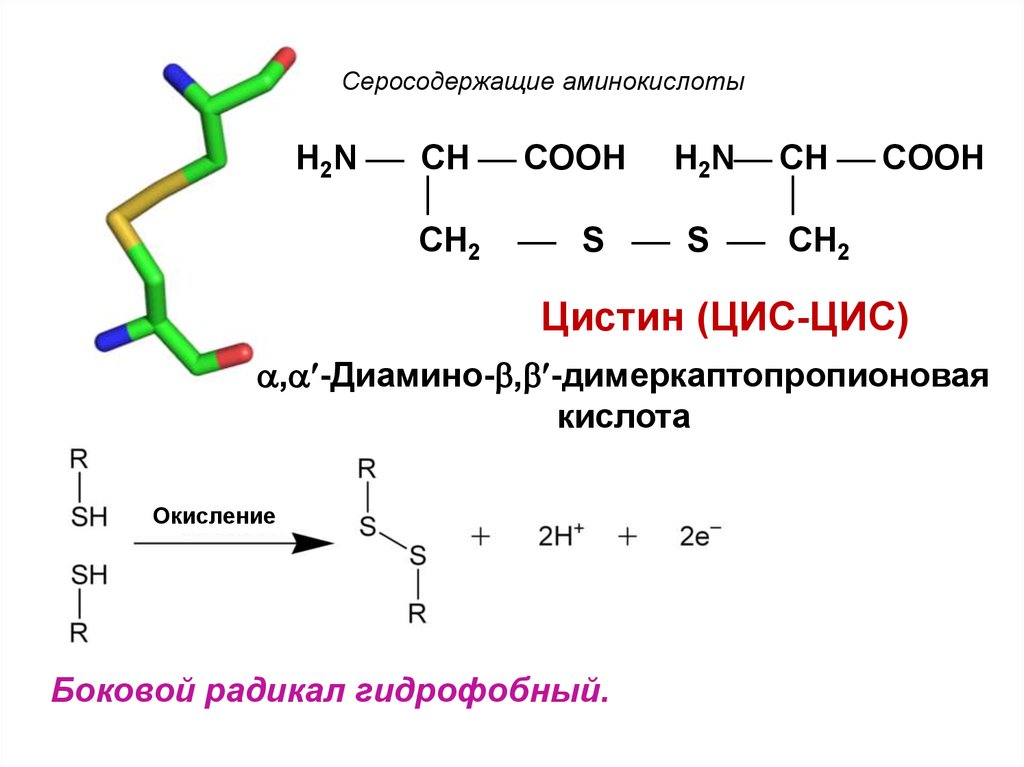 L-цистеин также является предшественником антиоксиданта глютатиона. Антиоксидантная активность L-метионина и метаболитов L-метионина, по-видимому, объясняет его возможную антигепатотоксическую активность. Недавние исследования показывают, что сам метионин обладает активностью удалять свободные радикалы благодаря сере, а также своей хелатирующей способности.
L-цистеин также является предшественником антиоксиданта глютатиона. Антиоксидантная активность L-метионина и метаболитов L-метионина, по-видимому, объясняет его возможную антигепатотоксическую активность. Недавние исследования показывают, что сам метионин обладает активностью удалять свободные радикалы благодаря сере, а также своей хелатирующей способности.
При метаболизме метионин входит в метиониновый цикл и превращается в S-аденозилметионин (SAMe). После пожертвования метильной группы SAMe гидролизуется до гомоцистеина, а затем либо переметилируется до метионина, либо пересыщается, что приводит к образованию цистеина, таурина и глутатиона. Как антиоксидант глутатион предотвращает повреждение печени свободными радикалами, а таурин играет роль в конъюгации желчных кислот. Считается, что неспособность поддерживать гомеостаз цикла метионина приводит к повреждению печени. У пациентов с алкогольным заболеванием печени часто наблюдается гиперметионинемия, которая, как полагают, обусловлена снижением метаболизма метионина до SAMe.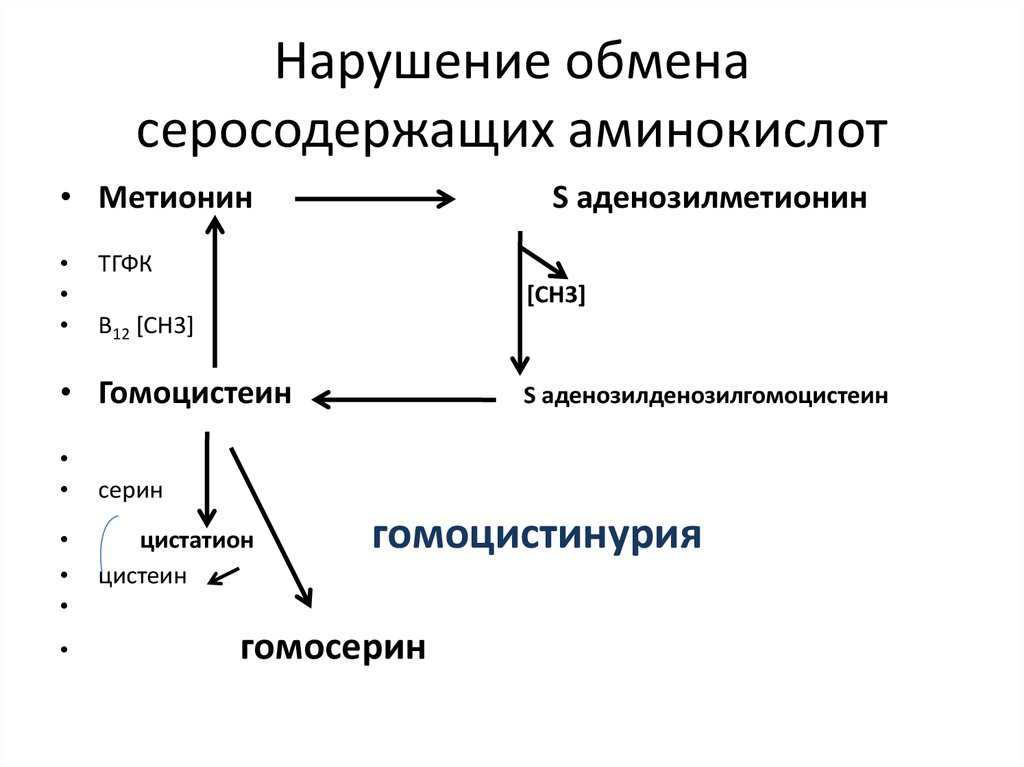 При низком уровне глутатиона в печени повышен риск повреждения свободными радикалами.
При низком уровне глутатиона в печени повышен риск повреждения свободными радикалами.
Токсичные уровни метионина могут быть скорректированы с помощью глицина. Глицин, по-видимому, усиливает разложение метионина путем транссульфурации, действуя в качестве рецептора для метильных групп. Избыток метионина конкурирует за глицин и ограничивает доступность глицина для других метаболических взаимодействий, таких как синтез глутатиона. При отравлении ацетаминофеном метионин, по-видимому, предотвращает повреждение печени и некроз, стимулируя синтез глутатиона. Токсичный метаболит ацетаминофена (N-ацетил-п-бензхинонимин) будет связываться с глутатионом вместо клеток печени.
Диетический метионин обычно не влияет на уровень гомоцистеина. Уровни могут повышаться и, возможно, вызывать гипергомоцистеинемию, если ферменты, используемые для метаболизма гомоцистеина, имеют дефекты и / или существует дефицит фолиевой кислоты, витаминов B6 или B12. Гипергомоцистеинемия может вызвать повреждение эндотелия и повысить риск развития сосудистых заболеваний.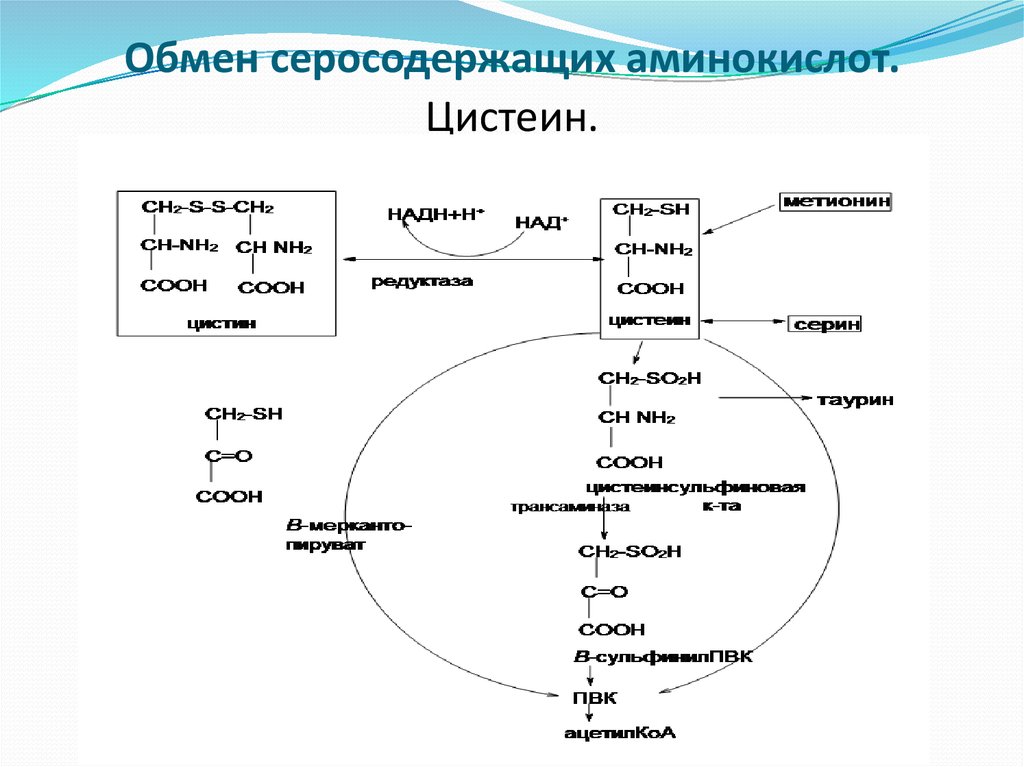 Гипергомоцистеинемия, которая не отвечает на витаминные добавки, иногда реагирует на диетическое ограничение метионина.
Гипергомоцистеинемия, которая не отвечает на витаминные добавки, иногда реагирует на диетическое ограничение метионина.
Метионин предотвращает побочные эффекты, вызванные длительным воздействием закиси азота. Токсичность закиси азота напоминает дефицит кобаламина. Предварительные данные свидетельствуют о том, что закись азота может избирательно нарушать функцию кобаламин-зависимой метионинсинтазы. Введение метионина до операции может предотвратить инактивацию метионинсинтазы и предотвратить инактивацию кобаламина у пациентов, подвергающихся наркозу закисью азота.
Известно, что многие аминокислоты стимулируют гормон роста. Существуют предварительные доказательства того, что метионин также усиливает секрецию базального гормона роста.
Метионин может действовать синергически с фолатом, снижая риск рака толстой кишки. Однако есть также свидетельства того, что высокое потребление метионина с солью и нитратами в рационе может увеличить риск рака желудка.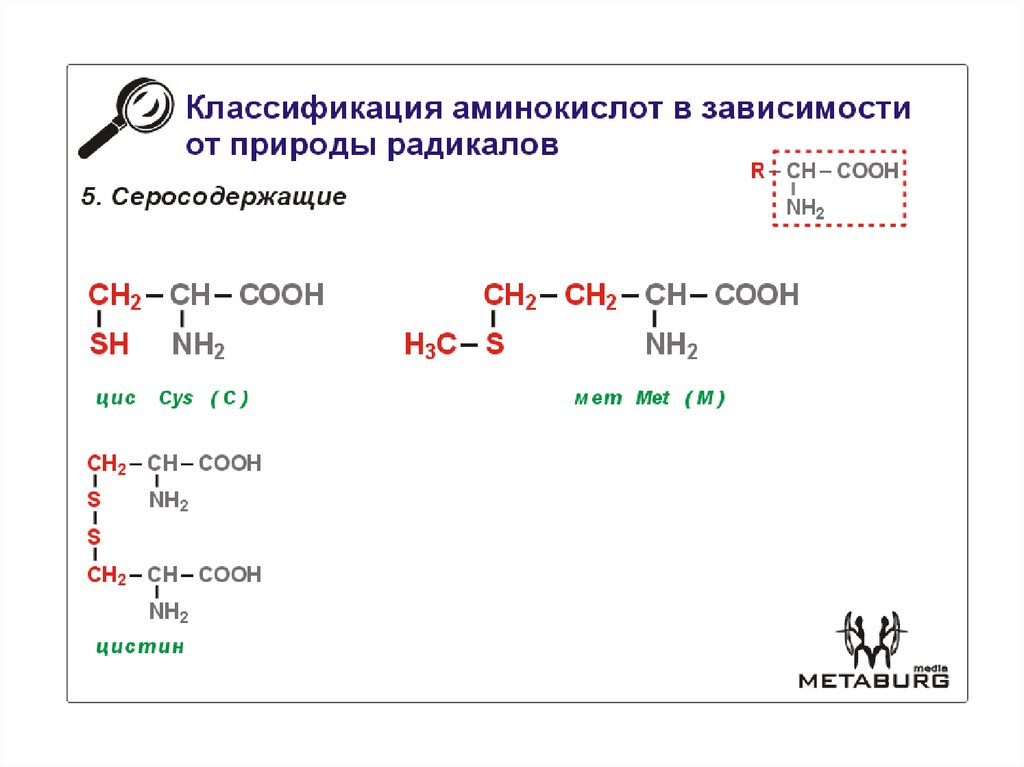 Уровень метилирования в опухоли намного выше, чем в нормальной ткани. Большинство опухолей зависят от экзогенного, предварительно образованного метионина для роста. Предварительные клинические данные свидетельствуют о том, что ограничение метионина в рационе у больных раком может ингибировать рост опухоли и улучшить результаты лечения рака.
Уровень метилирования в опухоли намного выше, чем в нормальной ткани. Большинство опухолей зависят от экзогенного, предварительно образованного метионина для роста. Предварительные клинические данные свидетельствуют о том, что ограничение метионина в рационе у больных раком может ингибировать рост опухоли и улучшить результаты лечения рака.
Токсикологические данные:
Острая токсичность LD50 при оральном применении — крыса — 3400 мг / кг
Серосодержащие аминокислоты и болезни человека
[1] Wagner C, Decha-Umphai W, Corbin J. Фосфорилирование модулирует активность глицин-N-метилтрансферазы, белка, связывающего фолиевую кислоту. Фосфорилирование in vitro ингибируется природным фолатным лигандом. J Биол Хим
1989;264(16):9638–42. [PubMed] [Google Scholar]
[2] Jencks DA, Mathews RG. Аллостерическое ингибирование метилентетрагидрофолатредуктазы аденозилметионином. Влияние аденозилметионина и НАДФН на равновесие между активной и неактивной формами фермента и на кинетику приближения к равновесию.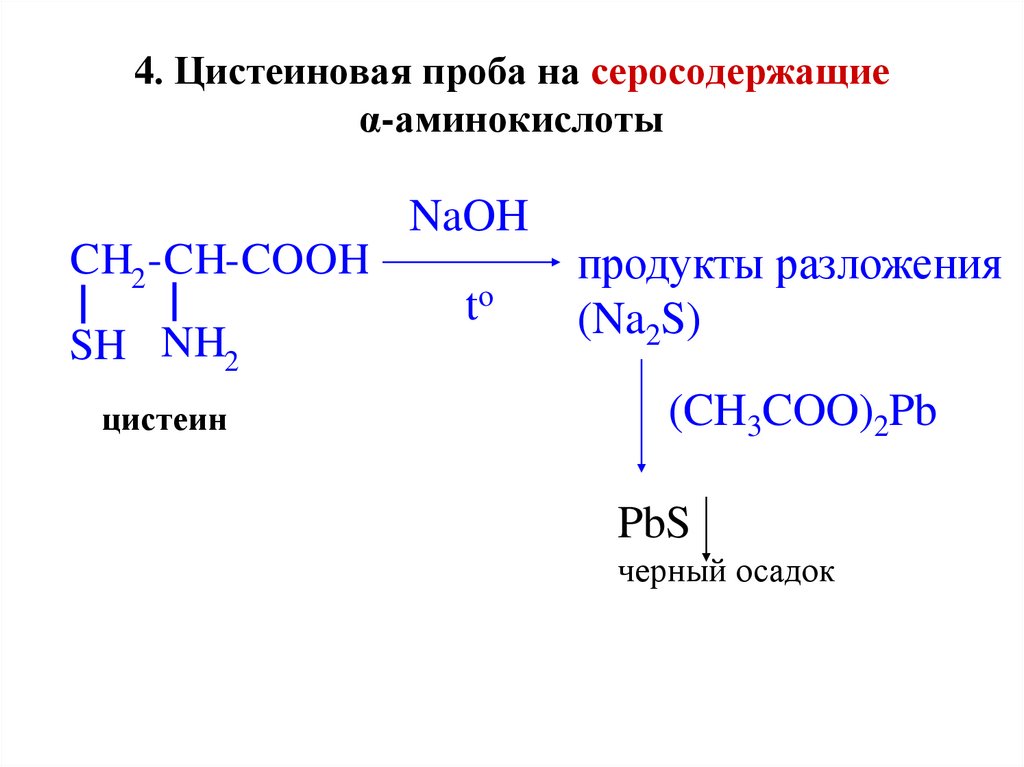 J Биол Хим
J Биол Хим
1987;262(6):2485–93. [PubMed] [Google Scholar]
[3] Wagner C, Briggs WT, Cook RJ. Ингибирование активности глицин-N-метилтрансферазы производными фолиевой кислоты: последствия для регуляции метаболизма метильных групп. Biochem Biophys Res Commun
1985;127(3):746–52. [PubMed] [Google Scholar]
[4] Kutzbach C, Stokstad EL. Ингибирование S-аденозилметионином метилентетрагидрофолатредуктазы в печени крыс по принципу обратной связи. Биохим Биофиз Акта
1967;139(1):217–20. [PubMed] [Академия Google]
[5] Townsend DM, Tew KD, Tapiero H. Важность глутатиона при заболеваниях человека. Биомед Фармаколог
2003; 57 (3–4): 145–55. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[6] Choi SW, Mason JB. Фолат и канцерогенез: комплексная схема. Джей Нутр
2000;130(2):129–32. [PubMed] [Google Scholar]
[7] Mason JB. Биомаркеры воздействия питательных веществ и состояния одноуглеродного (метилового) метаболизма. Джей Нутр
2003; 133 (Приложение 3): 941S–7S. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
[8] McCully KS. Сосудистая патология гомоцистеинемии: значение для патогенеза атеросклероза. Ам Джей Патол
1969;56(1):111–28. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[9] Clarke R, Daly L, Robinson K, Naughten E, Cahalane S, Fowler B, et al.
Гипергомоцистеинемия: независимый фактор риска сосудистых заболеваний. N Engl J Med
1991;324(17):1149–55. [PubMed] [Google Scholar]
[10] Канг С.С., Вонг П.В.К., Малинов М.Р. Гипергомоцист(е)инемия как фактор риска окклюзионных сосудистых заболеваний. Энн Рев Нутр
1992; 12: 279–98. [PubMed] [Google Scholar]
[11] Vollset SE, Refsum H, Tverdal A, Nygard O, Nordrehaug JE, Tell GS, et al.
Общий гомоцистеин плазмы и смертность от сердечно-сосудистых и несердечно-сосудистых заболеваний: исследование Hordaland Homocysteine Study. Am J Clin Nutr
2001;74(1):130–6. [PubMed] [Академия Google]
[12] МакКьюсик В.А. Наследственные заболевания соединительной ткани. 3-е изд.
Сент-Луис: CV Мосби; 1966. с. 155. [Google Scholar]
[13] Уббинк Дж. Б., Вермаак В. Дж., Ван дер Мерве А., Беккер П. Дж. Нутриционный статус витамина B-12, витамина B-6 и фолиевой кислоты у мужчин с гипергомоцистеинемией. Am J Clin Nutr
1993;57(1):47–53. [PubMed] [Google Scholar]
[14] Jacques PF, Selhub J, Bostom AG, Wilson PW, Rosenberg IH. Влияние обогащения фолиевой кислотой на концентрацию фолиевой кислоты в плазме и общий уровень гомоцистеина. N Engl J Med
1999;340(19):1449–54. [PubMed] [Google Scholar]
[15] Riddell LJ, Chisholm A, Williams S, Mann JI. Диетические стратегии для снижения концентрации гомоцистеина. Am J Clin Nutr
2000;71(6): 1448–54. [PubMed] [Google Scholar]
[16] Карсон Н.А. Нарушения обмена веществ обнаружены при обследовании умственно отсталых лиц в Северной Ирландии. Арка Дис Чайлд
1962; 37: 505–13. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[17] Gerritsen T, Vaughn JG, Waisman HA. Определение гомоцистина в моче. Biochem Biophys Res Commun
1962;9:493-6. [PubMed] [Google Scholar]
[18] Mudd SH, Skovby F, Levy HL, Pettigrew KD, Wilcken B, Pyeritz RE, et al.
Естественная история гомоцистинурии из-за дефицита цистатионин бета-синтазы. Am J Hum Genet
1985;37(1):1–31. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[19] Фаулер Б., Краус Дж., Пакман С., Розенберг Л.Е. Гомоцистинурия. Доказательства существования трех различных классов мутантов цистатионин-бета-синтазы в культивируемых фибробластах. Джей Клин Инвест
1978;61(3):645–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[20] Сковби Ф., Красиков Н., Франке У. Назначение гена цистатионин-бета-синтазы хромосоме 21 человека в гибридах соматических клеток. Хум Жене
1984;65(3):291–4. [PubMed] [Google Scholar]
[21] Munke M, Kraus JP, Ohura T, Francke U. Ген цистатионин-бета-синтазы (CBS) картируется в субтеломерной области на хромосоме 21q человека и в проксимальной мышиной хромосоме 17. Am J Hum Genet
1988;42(4):550–9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[22] Kraus JP, Oliveriusova J, Sokolova J, Kraus E, Vlcek C, de Franchis R, et al.
Ген цистатионинбета-синтазы (CBS) человека: полная последовательность, альтернативный сплайсинг и полиморфизмы. Геномика
1998;52(3):312–24. [PubMed] [Google Scholar]
[23] Kraus JP, Janosik M, Kozich V, Mandell R, Shih V, Sperandeo MP, et al.
Мутации цистатионин-бета-синтазы при гомоцистинурии. Хум Мутат
1999;13(5):362–75. [PubMed] [Google Scholar]
[24] Kraus JP. Комроверская лекция. Молекулярные основы экспрессии фенотипа при гомоцистинурии. J Наследовать Метаб Дис
1994;17(4):383–90. [PubMed] [Google Scholar]
[25] Abbott MH, Folstein SE, Abbey H, Pyeritz RE. Психиатрические проявления гомоцистинурии из-за дефицита цистатионин бета-синтазы: распространенность, естественное течение и связь с неврологическими нарушениями и реакцией на витамин B6. Am J Med Genet
1987; 26(4):959–69. [PubMed] [Google Scholar]
[26] Frimpter GW. Цистатионинурия: характер порока. Наука
1965 год; 149 (688): 1095–1096. [PubMed] [Google Scholar]
[27] Вонг Л.Т., Хардвик Д. Ф., Эпплгарт Д.А., Дэвидсон А.Г. Обзор программы метаболического скрининга детской больницы, Ванкувер, Британская Колумбия. 1971–1977 гг. Клин Биохим
1979; 12(5):167–72. [PubMed] [Google Scholar]
[28] Smith AJ, Strang LB. Врожденная ошибка метаболизма с выделением с мочой альфа-гидроксимасляной кислоты и фенилпировиноградной кислоты. Arch Dis Child
1958;33:109–13. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[29] Tapiero H, Tew KD, Gate L, Machover D. Профилактика патологий, связанных с окислительным стрессом и дефицитом питания: дефицит фолиевой кислоты и потребности. Биомед Фармаколог
2001; 55: 381–90. [PubMed] [Google Scholar]
[30] Лухби А.Л., Куперман Дж.М., Пеши-Бурель А. Новая врожденная ошибка метаболизма: реагирующая на фолиевую кислоту мегалобластная анемия, атаксия, умственная отсталость и судороги. Дж. Педиат
1965;67:1052 [Google Академия]
[31] Ланцковский П., Эрландсон М.Е., Безан А.И. Изолированный дефект всасывания фолиевой кислоты, связанный с умственной отсталостью и кальцинозом головного мозга. Кровь
1969;34(4):452–65. [PubMed] [Google Scholar]
[32] Wollaston WH. Кистозный оксид, новый вид мочевого конкремента. Фил Транс Рой Сок Лонд
1810; 100: 223–30. [Google Scholar]
[33] Calonge MJ, Gasparini P, Chillaron J, Chillon M, Gallucci M, Rou-saud F, et al.
Цистинурия, вызванная мутациями rBAT, гена, участвующего в транспорте цистина. Нат Жене
1994;6(4):420–5. [PubMed] [Google Scholar]
[34] Lee WS, Wells RG, Sabbag RV, Mohandas TK, Hediger MA. Клонирование и хромосомная локализация кДНК почки человека, участвующей в транспорте цистина, двухосновных и нейтральных аминокислот. Джей Клин Инвест
1993 год; 91 (5): 1959–63. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[35] Feliubadalo L, Font M, Purroy J, Rousaud F, Estivill X, Nunes V, et al.
Цистинурия не типа I, вызванная мутациями в SLC7A9, кодирующем субъединицу (bo+AT) rBAT. Международный консорциум по цистинурии. Нат Жене
1999;23(1):52–7. [PubMed] [Google Scholar]
[36] Leclerc D, Wilson A, Dumas R, Gafuik C, Song D, Watkins D, et al.
Клонирование и картирование кДНК метионинсинтазы редуктазы, дефектного флавопротеина у пациентов с гомоцистинурией. Proc Natl Acad Sci USA
1998;95(6):3059–64. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[37] Wilson A, Leclerc D, Rosenblatt DS, Gravel RA. Молекулярная основа дефицита метионинсинтазы редуктазы у пациентов, принадлежащих к группе комплементарных cblE нарушений метаболизма фолиевой кислоты/кобаламина. Хум Мол Жене
1999;8(11):2009–16. [PubMed] [Google Scholar]
[38] Wilson A, Platt R, Wu Q, Leclerc D, Christensen B, Yang H, et al.
Распространенный вариант редуктазы метионинсинтазы в сочетании с низким содержанием кобаламина (витамин B12) увеличивает риск расщепления позвоночника. Молек Генет Метаб
1999; 67: 317–23. [PubMed] [Google Scholar]
[39] Hobbs CA, Sherman SL, Yi P, Hopkins SE, Torfs CP, Hine RJ, et al.
Полиморфизмы генов, участвующих в метаболизме фолиевой кислоты, как факторы материнского риска синдрома Дауна. Am J Hum Genet
2000;67(3):623–30. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[40] James SJ, Pogribna M, Pogribny IP, Melnyk S, Hine RJ, Gibson JB, et al.
Аномальный метаболизм фолиевой кислоты и мутация в гене метилентетрагидрофолатредуктазы могут быть факторами риска развития синдрома Дауна у матери. Am J Clin Nutr
1999;70(4):495–501. [PubMed] [Google Scholar]
[41] Дулин М.Т., Барбо С., Макдоннелл М., Хёсс К., Уайтхед А.С., Митчелл Л.Е. Материнские генетические эффекты, оказываемые генами, участвующими в реметилировании гомоцистеина, влияют на риск расщепления позвоночника. Am J Hum Genet
2002;71(5):1222–6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[42] Гоган Д.Дж., Барбо С., Клюйтманс Л.А., Уайтхед А.С. Гены метилентетрагидрофолатредуктазы (MTHFR) человека и мыши: геномная организация, структура мРНК и связь с геном CLCN6. Ген
2000;257(2):279–89. [PubMed] [Google Scholar]
[43] Sibani S, Christensen B, O’Ferrall E, Saadi I, Hiou-Tim F, Rosenblatt DS, et al.
Характеристика шести новых мутаций в гене метилентетрагидрофолатредуктазы (MTHFR) у пациентов с гомоцистинурией. Хум Мутат
2000;15(3):280–7. [PubMed] [Академия Google]
[44] Розен Р
Молекулярная генетика дефицита метилентетрагидрофолатредуктазы. J Наследовать Метаб Дис
1996;19(5):589–94. [PubMed] [Google Scholar]
[45] Goyette P, Sumner JS, Milos R, Duncan AM, Rosenblatt DS, Matthews RG, et al.
Метилентетрагидрофолатредуктаза человека: выделение кДНК, картирование и идентификация мутаций. Нат Жене
1994 год; 7(2):195–200. [PubMed] [Google Scholar]
[46] Frosst P, Blom HJ, Milos R, Goyette P, Sheppard CA, Matthews RG, et al.
Кандидатный генетический фактор риска сосудистых заболеваний: распространенная мутация метилентетрагидрофолатредуктазы. Нат Жене
1995;10(1):111–3. [PubMed] [Google Scholar]
[47] Schneider JA, Rees DC, Liu YT, Clegg JB. Всемирное распространение распространенной мутации метилентетрагидрофолатредуктазы. Am J Hum Genet
1998;62(5):1258–60. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[48] McAndrew PE, Brandt JT, Pearl DK, Prior TW. Распространенность гена термолабильной метилентетрагидрофолатредуктазы у афроамериканцев. Тромб Res
1996;83(2):195–8. [PubMed] [Google Scholar]
[49] Bragley PJ, Selhub J. Распространенная мутация в гене метилентетрагидрофолатредуктазы связана с накоплением формилированных тетрагидрофолатов в эритроцитах. Proc Natl Acad Sci USA
1998;95:13217–20. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[50] Friso S, Choi SW, Girelli D, Mason JB, Dolnikowski GG, Bagley PJ, et al.
Распространенная мутация в гене 5,10-метилентетрагидрофолатредуктазы влияет на метилирование геномной ДНК посредством взаимодействия с фолатным статусом. Proc Natl Acad Sci USA
2002;99(8):5606–11. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[51] Jacques PF, Bostom AG, Williams RR, Ellison RC, Eckfeldt JH, Rosenberg IH, et al.
Связь между фолатным статусом, распространенной мутацией метилентетрагидрофолатредуктазы и концентрацией гомоцистеина в плазме. Тираж
1996;93(1):7–9. [PubMed] [Google Scholar]
[52] Kluijtmans LA, van den Heuvel LP, Boers GH, Frosst P, Stevens EM, van Oost BA, et al.
Молекулярно-генетический анализ при легкой гипергомоцистеинемии: распространенная мутация в гене метилентетрагидрофолатредуктазы является генетическим фактором риска сердечно-сосудистых заболеваний. Am J Hum Genet
1996;58(1):35–41. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[53] Klerk M, Verhoef P, Clarke R, Blom HJ, Kok FJ, Schouten EG. MTHFR 677C → полиморфизм T и риск ишемической болезни сердца: метаанализ. ДЖАМА
2002;288(16):2023–31. [PubMed] [Академия Google]
[54] Christensen B, Arbor L, Tran P, Leclerc D, Sabbaghian N, Platt R, et al.
Генетический полиморфизм метилентетрагидрофолатредуктазы и метионинсинтазы, уровни фолиевой кислоты в эритроцитах и риск дефектов нервной трубки. Am J Med Genet
1999;84(2):151–7. [PubMed] [Google Scholar]
[55] Ou CY, Stevenson RE, Brown VK, Schwartz CE, Allen WP, Khoury MJ, et al.
5,10 Генетический полиморфизм метилентетрагидрофолатредуктазы как фактор риска дефектов нервной трубки. Am J Med Genet
1996;63(4):610–4. [PubMed] [Google Scholar]
[56] Mornet E, Muller F, Lenvoise-Furet A, Delezoide AL, Col JY, Simon-Bouy B, et al.
Скрининг мутации C677T в гене метилентетрагидрофолатредуктазы у французских пациентов с дефектами нервной трубки. Хум Жене
1997; 100 (5–6): 512–4. [PubMed] [Google Scholar]
[57] Speer MC, Worley G, Mackey JF, Melvin E, Oakes WJ, George TM. Термолабильный вариант метилентетрагидрофолатредуктазы (MTHFR) не является основным фактором риска дефекта нервной трубки у американских европеоидов. Совместная группа NTD. нейрогенетика
1997; 1(2):149–50. [PubMed] [Google Scholar]
[58] Мотульский А.Г. Экогенетика питания: артериосклеротическое заболевание сосудов, связанное с гомоцистеином, дефекты нервной трубки и фолиевая кислота. Am J Hum Genet
1996;58(1):17–20. [PMC free article] [PubMed] [Google Scholar]
[59] Pietrzyk JJ, Bik-Multanowski M, Sanak M, Twardowska M. Полиморфизмы генов 5,10-метилентетрагидрофолата и метионинсинтазы редуктазы как независимые факторы риска расщепление позвоночника. J Appl Genet
2003;44(1):111–3. [PubMed] [Академия Google]
[60] Погрибна М., Мельник С., Погрибный И., Чанго А., Йи П., Джеймс С.Дж. Метаболизм гомоцистеина у детей с синдромом Дауна: модуляция in vitro. Am J Hum Genet
2001;69(1):88–95. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[61] O’Leary VB, Parle-McDermott A, Molloy AM, Kirke PN, Johnson Z, Conley M, et al.
Полиморфизм MTRR и MTHFR: связь с синдромом Дауна?
Am J Med Genet
2002;107(2):151–5. [PubMed] [Google Scholar]
[62] Wisniewski KE, Wisniewski HM, Wen GY. Возникновение нейропатологических изменений и деменции болезни Альцгеймера при синдроме Дауна. Энн Нейрол
1985;17(3):278–82. [PubMed] [Google Scholar]
[63] Religa D, Styczynska M, Peplonska B, Gabryelewicz T, Pfeffer A, Chodakowska M, et al.
Гомоцистеин, аполипопротеин Е и метилентетрагидрофолатредуктаза при болезни Альцгеймера и легких когнитивных нарушениях. Дементное гериатрическое когнитивное расстройство
2003;16(2):64–70. [PubMed] [Google Scholar]
[64] Хоффман Р. М. Зависимость от метионина в раковых клетках — обзор. В пробирке
1982;18(5):421–8. [PubMed] [Google Scholar]
[65] Хоффман Р.М. Измененный метаболизм метионина и трансметилирование при раке. Противораковый Рез
1985;5(1):1–30. [PubMed] [Google Scholar]
[66] Liteplo RG, Hipwell SE, Rosenblatt DS, Sillaots S, Lue-Shing H. Изменения в метаболизме кобаламина связаны с измененной ауксотрофией метионина в клетках меланомы человека с высоким автономным ростом. J Cell Физиол
1991;149(2):332–38. [PubMed] [Google Scholar]
[67] Tautt JW, Anuszewska EL, Koziorowska JH. Метиониновая регуляция N-5-метилтетрагидрофолата: гомоцистеинметилтрансфераза и ее влияние на рост и синтез белка в нормальных, неопластических и трансформированных клетках в культуре. J Natl Институт рака
1982;69(1):9–14. [PubMed] [Google Scholar]
[68] Fiskerstrand T, Christensen B, Tysnes OB, Ueland PM, Refsum H. Развитие и реверсия зависимости от метионина в клеточной линии глиомы человека: связь с реметилированием гомоцистеина и статусом кобаламина. Рак Рез
1994;54(18):4899–906. [PubMed] [Google Scholar]
[69] Тан Б., Ли Ю.Н., Крюгер В.Д. Дефекты метилтиоаденозинфосфорилазы связаны с метионин-зависимым ростом опухолевых клеток, но не ответственны за него. Рак Рез
2000;60(19): 5543–7. [PubMed] [Google Scholar]
[70] Мацуо К., Хамадзима Н., Хираи Т., Като Т., Иноуэ М., Такезаки Т. и др.
Полиморфизм гена метионинсинтазы редуктазы A66G связан с риском колоректального рака. Asian Pac J Cancer Prev
2002;3(4): 353–9. [PubMed] [Google Scholar]
[71] Paz MF, Avila S, Fraga MF, Pollan M, Capella G, Peinado MA, et al.
Варианты зародышевой линии в генах метаболизма метильных групп и восприимчивость к метилированию ДНК в нормальных тканях и первичных опухолях человека. Рак Рез
2002;62(15):4519–24. [PubMed] [Google Scholar]
[72] Skibola CF, Smith MT, Kane E, Roman E, Rollinson S, Cart-wright RA, et al.
Полиморфизмы гена метилентетрагидрофолатредуктазы связаны с предрасположенностью к острому лейкозу у взрослых. Proc Natl Acad Sci USA
1999;96(22):12810–5. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[73] Wiemels JL, Smith RN, Taylor GM, Eden OB, Alexander FE, Greaves MF. Полиморфизмы метилентетрагидрофолатредуктазы (MTHFR) и риск молекулярно определенных подтипов острого лейкоза у детей. Proc Natl Acad Sci USA
2001;98(7):4004–9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Серосодержащие аминокислоты: обзор
Обзор
. 2006 г., июнь; 136 (6 доп.): 1636S-1640S.
дои: 10.1093/jn/136.6.1636S.
Джон Т. Броснан
1
, Маргарет Э. Броснан
принадлежность
- 1 Кафедра биохимии, Мемориальный университет Ньюфаундленда, Сент-Джонс, Нидерланды, Канада A1B 3X9. [email protected]
PMID:
16702333
DOI:
10.
1093/ин/136.6.1636С
Обзор
John T Brosnan et al.
Дж Нутр.
2006 9 июня0003
. 2006 г., июнь; 136 (6 доп.): 1636S-1640S.
дои: 10.1093/jn/136.6.1636S.
Авторы
Джон Т. Броснан
1
, Маргарет Э. Броснан
принадлежность
- 1 Кафедра биохимии, Мемориальный университет Ньюфаундленда, Сент-Джонс, Нидерланды, Канада A1B 3X9. [email protected]
PMID:
16702333
DOI:
10.
1093/ин/136.6.1636С
Абстрактный
Метионин, цистеин, гомоцистеин и таурин являются 4 распространенными серосодержащими аминокислотами, но только первые 2 входят в состав белков. Сера принадлежит к той же группе в периодической таблице, что и кислород, но гораздо менее электроотрицательна. Это различие объясняет некоторые отличительные свойства серосодержащих аминокислот. Метионин является инициирующей аминокислотой в синтезе практически всех эукариотических белков; N-формилметионин выполняет ту же функцию у прокариот. В белках многие остатки метионина скрыты в гидрофобном ядре, но некоторые из них, выставленные напоказ, подвержены окислительному повреждению. Цистеин, благодаря своей способности образовывать дисульфидные связи, играет решающую роль в структуре белка и в путях его сворачивания. Метаболизм метионина начинается с его активации до S-аденозилметионина. Это чрезвычайно универсальный кофактор, играющий роль в переносе метильной группы, переносе 5′-дезоксиаденозильной группы, синтезе полиаминов, синтезе этилена в растениях и многих других. У животных большая часть S-аденозилметионина используется в реакциях метилирования. S-аденозилгомоцистеин, который является продуктом этих метилтрансфераз, дает гомоцистеин. Гомоцистеин может реметилироваться в метионин или превращаться в цистеин по пути транссульфирования. Метионин также может метаболизироваться по пути трансаминирования. Этот путь, который важен только при высоких концентрациях метионина, производит ряд токсичных конечных продуктов. Цистеин может быть преобразован в такие важные продукты, как глутатион и таурин. Таурин присутствует во многих тканях в более высоких концентрациях, чем любые другие аминокислоты. Это важное питательное вещество для кошек.
Похожие статьи
Метаболизм серосодержащих аминокислот.
Стипанюк М.Х.
Стипанюк МХ.
Анну Рев Нутр. 1986; 6:179-209. doi: 10.1146/annurev.nu.06.070186.001143.
Анну Рев Нутр. 1986 год.PMID: 3524616
Обзор.
Работа с метиониновой/гомоцистеиновой серой: метаболизм цистеина в таурин и неорганическую серу.
Стипанюк М.Х., Уэки И.
Стипанюк М.Х. и соавт.
J Наследовать Metab Dis. 2011 фев; 34 (1): 17-32. doi: 10.1007/s10545-009-9006-9. Epub 2010 17 февраля.
J Наследовать Metab Dis. 2011.PMID: 20162368
Бесплатная статья ЧВК.Обзор.
Влияние добавок бетаина на метаболизм серосодержащих аминокислот в печени у мышей.
Ким С.К., Ким Ю.К.
Ким С.К. и др.
J Гепатол. 2005 июнь; 42 (6): 907-13. doi: 10.1016/j.jhep.2005.01.017. Epub 2005 1 апр.
J Гепатол. 2005.PMID: 15885362
Печеночный метаболизм аминокислот серы у мышей db/db.
Yun KU, Ryu CS, Lee JY, Noh JR, Lee CH, Lee HS, Kang JS, Park SK, Kim BH, Kim SK.
Юн Ку и др.
Пищевая химическая токсикол. 2013 март; 53:180-6. doi: 10.1016/j.fct.2012.11.046. Epub 2012 5 декабря.
Пищевая химическая токсикол. 2013.PMID: 23220616
Метаболизм аминокислот серы у крыс Цукера с диабетом и ожирением.
Квак Х.К., Ким Ю.М., О С.Дж., Ким С.К.
Квак Х.К. и др.
Биохим Фармакол. 2015 1 августа; 96 (3): 256-66. doi: 10.1016/j.bcp.2015.05.014. Epub 2015 3 июня.
Биохим Фармакол. 2015.PMID: 26047850
Посмотреть все похожие статьи
Цитируется
Влияние замены таурина метионином и дополнительной добавки таурина на продуктивность и антиоксидантную способность кур-несушек.
Юань Х, Лю Ю, Чен Ю, Цзяо Х, Чжао Дж, Ван Х, Чжоу Ю, Линь Х.
Юань X и др.
Poult Sci. 2022 г., 15 декабря; 102(3):102426. doi: 10.1016/j.psj.2022.102426. Онлайн перед печатью.
Poult Sci. 2022.PMID: 36587450
Бесплатная статья ЧВК.Серосодержащие метаболиты из морских и наземных источников грибов: происхождение, структура и биологическая активность.
Лю З., Ли М., Ван С., Хуан Х., Чжан В.
Лю Зи и др.
Мар Наркотики. 2022 7 декабря; 20 (12): 765. doi: 10.3390/md20120765.
Мар Наркотики. 2022.PMID: 36547912
Бесплатная статья ЧВК.Обзор.
Содержание кофеина, D-глюкуронолактона и таурина в энергетических напитках: воздействие и оценка риска.
Рубио С., Камара М., Хинер Р.
М., Гонсалес-Муньос М.Х., Лопес-Гарсия Э., Моралес Ф.Х., Морено-Аррибас М.В., Портильо М.П., Бетанкур Э.
Рубио С. и др.
Питательные вещества. 2022 1 декабря; 14 (23): 5103. дои: 10.3390/nu14235103.
Питательные вещества. 2022.PMID: 36501132
Бесплатная статья ЧВК.Новые комплексы Eu(III), Tb(III), Gd(III) и Sm(III) на основе 2,2′-бипиридина, присоединенные к ТЕМПО: синтез, фотофизические исследования и тестирование возможностей биовизуализации на основе фотолюминесценции.
Словеснова Н.В., Минин А.С., Белоусова А.В., Устюгов А.А., Чапров К.Д., Криночкин А.П., Валиева М.И., Штайц Ю.К., Старновская Е.С., Никонов И.Л., Цмокалюк А.Н., Ким Г.А., Сантра С., Копчук Д.С., Носова Е.В., Зырянов Г.В.
Словеснова Н.В., и соавт.
Молекулы. 2022 1 декабря; 27 (23): 8414. doi: 10,3390/молекулы27238414.
Молекулы. 2022.PMID: 36500504
Бесплатная статья ЧВК.