Сложный фермент не содержит: — строение ферментов — Биохимия
— строение ферментов — Биохимия
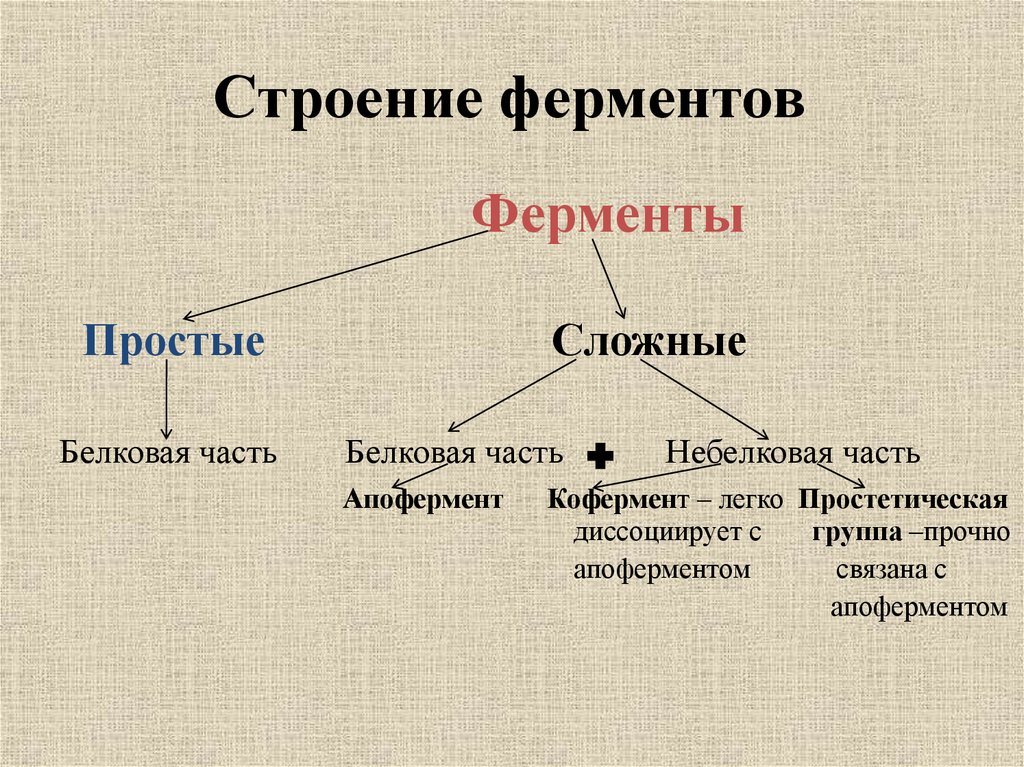
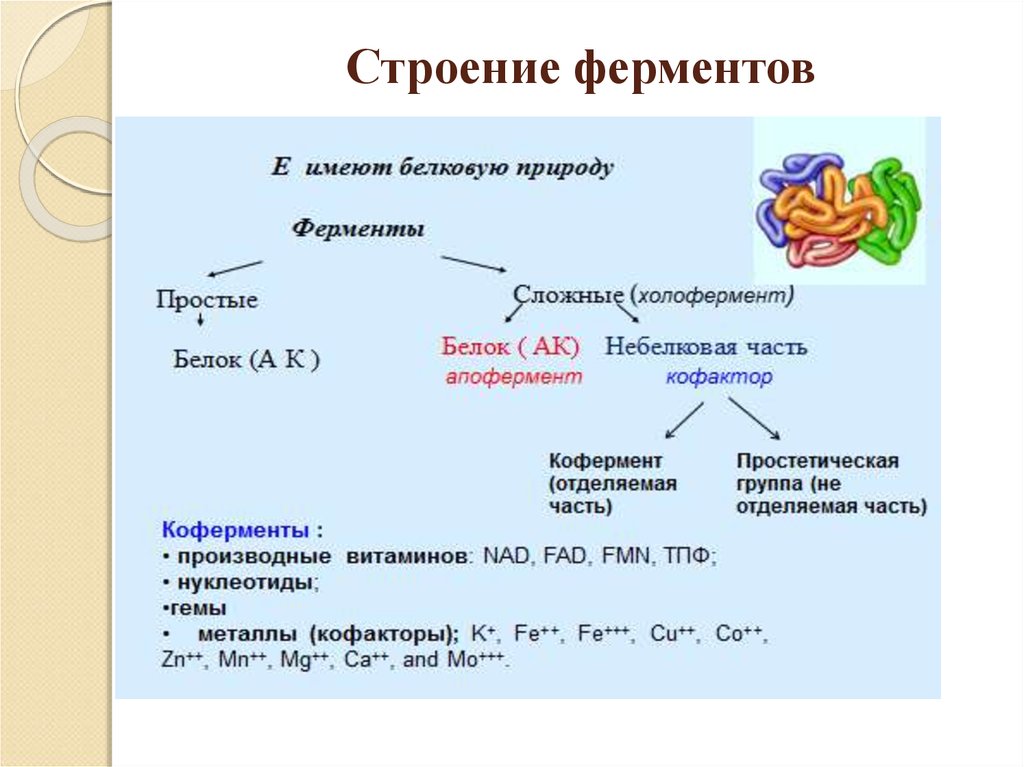
Давно выяснено, что все ферменты являются белками и обладают всеми свойствами белков. Поэтому, подобно белкам, ферменты делятся на простые и сложные.
Простые ферменты состоят только из аминокислот – например, пепсин , трипсин, лизоцим.
Сложные ферменты (холоферменты) имеют в своем составе белковую часть, состоящую из аминокислот – апофермент, и небелковую часть – кофактор. Примером сложных ферментов являются сукцинатдегидрогеназа (содержит ФАД), аминотрансферазы (содержат пиридоксальфосфат), различные пероксидазы (содержат гем), лактатдегидрогеназа (содержит Zn2+), амилаза (содержит Ca2+).
Кофактор, в свою очередь, может называться коферментом (НАД+, НАДФ+, ФМН, ФАД, биотин) или простетической группой (гем, олигосахариды, ионы металлов Fe2+, Mg2+, Ca2+, Zn2+).
Деление на коферменты и простетические группы не всегда однозначно:
• если связь кофактора с белком прочная, то в этом случае говорят о наличии простетической группы,
• но если в качестве кофактора выступает производное витамина – то его называют коферментом, независимо от прочности связи.
Для осуществления катализа необходим полноценный комплекс апобелка и кофактора, по отдельности катализ они осуществить не могут. Кофактор входит в состав активного центра, участвует в связывании субстрата или в его превращении.
Как многие белки, ферменты могут быть мономерами, т.е. состоять из одной субъединицы, и полимерами, состоящими из нескольких субъединиц.
Структурно-функциональная организация ферментов
В составе фермента выделяют области, выполняющие различную функцию:
1. Активный центр – комбинация аминокислотных остатков (обычно 12-16), обеспечивающая непосредственное связывание с молекулой субстрата и осуществляющая катализ. Аминокислотные радикалы в активном центре могут находиться в любом сочетании, при этом рядом располагаются аминокислоты, значительно удаленные друг от друга в линейной цепи. В активном центре выделяют два участка:
Аминокислотные радикалы в активном центре могут находиться в любом сочетании, при этом рядом располагаются аминокислоты, значительно удаленные друг от друга в линейной цепи. В активном центре выделяют два участка:
- якорный (контактный, связывающий) – отвечает за связывание и ориентацию субстрата в активном центре,
- каталитический – непосредственно отвечает за осуществление реакции.
Схема строения ферментов
У ферментов, имеющих в своем составе несколько мономеров, может быть несколько активных центров по числу субъединиц. Также две и более субъединицы могут формировать один активный центр.
У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора. Например, в реакции превращения пировиноградной кислоты (пируват) в молочную кислоту (лактат) сначала к апоферменту лактатдегидрогеназы присоединяется НАД, формируется активный центр, и только потом входит пируват.
Схема формирования сложного фермента
2. Аллостерический центр (allos – чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы, называемой активатором или ингибитором (или эффектором, модулятором, регулятором), вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции.
Аллостерический центр (allos – чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы, называемой активатором или ингибитором (или эффектором, модулятором, регулятором), вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции.
Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся в разных субъединицах.
Схема строения аллостерического фермента
В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество (см «Регуляция активности ферментов»).
Изоферменты
Изоферменты – это молекулярные формы одного и того же фермента, возникшие в результате небольших генетических различий в первичной структуре фермента, но катализирующие одну и ту же реакцию. Изоферменты отличаются сродством к субстрату, максимальной скоростью катализируемой реакции, чувствительностью к ингибиторам и активаторам, условиями работы (оптимум pH и температуры).
Как правило, изоферменты имеют четвертичную структуру, т.е. состоят из двух или более субъединиц. Например, димерный фермент креатинкиназа (роль КК) представлен тремя изоферментными формами, составленными из двух типов субъединиц: M (англ. muscle – мышца) и B (англ. brain – мозг). Креатинкиназа-1 (КК-1) состоит из субъединиц типа B и локализуется в головном мозге, креатинкиназа-2 (КК-2) – по одной М- и В-субъединице, наиболее активна в миокарде, креатинкиназа-3 (КК-3) содержит две М-субъединицы, специфична для скелетной мышцы. Определение активности разных изоферментов КК в сыворотке крови имеет клинико-диагностическое значение.
Необходимо понимать, что хотя в головном мозге активность креатинкиназы ВВ составляет все 100%, ее высокая активность обнаружена также в стенке желудка и почках. В скелетных мышцах имеется только креатинкиназа ММ (100%), но довольно высока ее активность также в печени и в сердце. Креатинкиназа МВ в сердце составляет лишь 22% от общей креатинкиназной активности, в большем количестве здесь имеется КК ММ, но зато в других тканях креатинкиназы МВ практически нет.
Изоферменты креатинкиназы
|
Изоферменты лактатдегидрогеназы
|
Также существует пять изоферментов лактатдегидрогеназы (роль ЛДГ) – фермента, участвующего в обмене глюкозы. Отличия между ними заключаются в разном соотношении субъединиц Н (англ. heart – сердце) и М (англ. muscle – мышца). Лактатдегидрогеназы типов 1 (Н4) и 2 (H3M1) присутствуют в тканях с аэробным обменом (миокард, мозг, корковый слой почек), обладают высоким сродством к молочной кислоте (лактату) и превращают его в пируват. Изоферменты ЛДГ-4 (H1M3) и ЛДГ-5 (М4) находятся в тканях, склонных к анаэробному обмену (печень, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2). Определение активности разных изоферментов ЛДГ в сыворотке крови имеет клинико-диагностическое значение.
В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2). Определение активности разных изоферментов ЛДГ в сыворотке крови имеет клинико-диагностическое значение.
Еще одним примером изоферментов является группа гексокиназ, которые присоединяют фосфатную группу к моносахаридам гексозам и вовлекают их в реакции клеточного метаболизма. Из четырех изоферментов выделяется гексокиназа IV (глюкокиназа), которая отличается от остальных изоферментов высокой специфичностью к глюкозе, низким сродством к ней и нечувствительностью к ингибированию продуктом реакции.
Мультиферментные комплексы
В мультиферментном комплексе несколько ферментов прочно связаны между собой в единый комплекс и осуществляют ряд последовательных реакций, в которых продукт реакции непосредственно передается на следующий фермент и является только его субстратом. Возникает туннельный эффект, т.е. субстрат попадает в созданный ферментами «туннель». В результате промежуточные метаболиты избегают контакта с окружающей средой, снижается время их перехода к следующему активному центру и значительно ускоряется скорость реакции.
Возникает туннельный эффект, т.е. субстрат попадает в созданный ферментами «туннель». В результате промежуточные метаболиты избегают контакта с окружающей средой, снижается время их перехода к следующему активному центру и значительно ускоряется скорость реакции.
Строение мульферментного комплекса
Например,
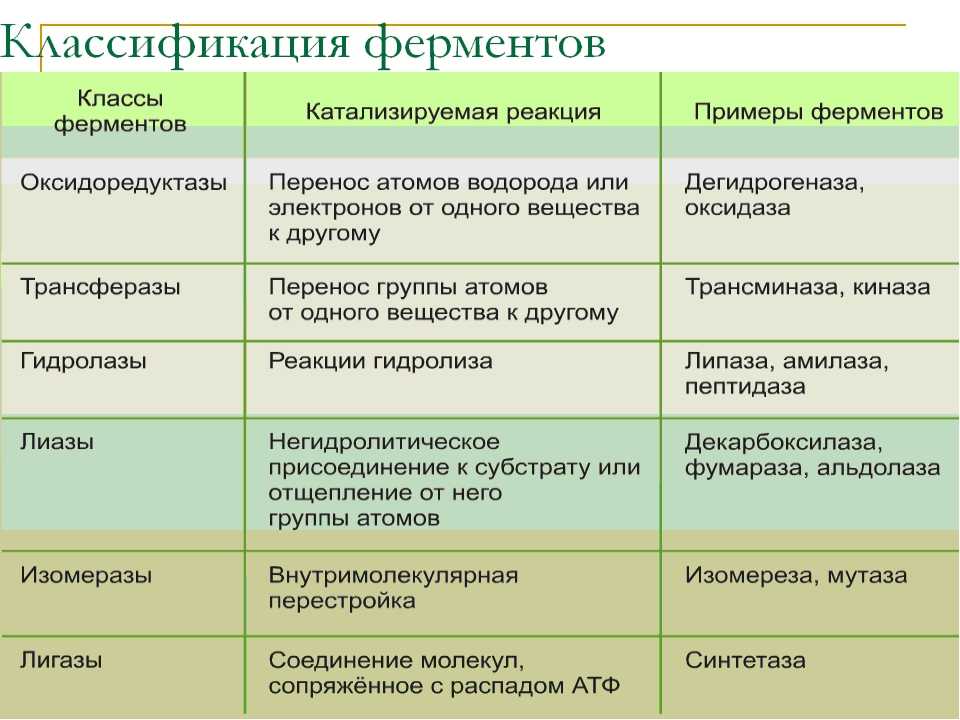
- пируватдегидрогеназный комплекс (пируватдегидрогеназа), превращающий пируват в ацетил-SКоА,
- α-кетоглутаратдегидрогеназный комплекс (в цикле трикарбоновых кислот) превращающий α-кетоглутарат в сукцинил-SКоА,
- комплекс под названием «синтаза жирных кислот» (или пальмитатсинтаза), синтезирующий пальмитиновую кислоту
- карбамоилфосфатсинтетаза, первый фермент синтеза мочевины в печени и синтеза пиримидиновых оснований
Абзимы
Абзимами называются антитела, имеющие каталитическую функцию (англ. abzymes, antibodies as enzymes) и катализирующие конкретные реакции.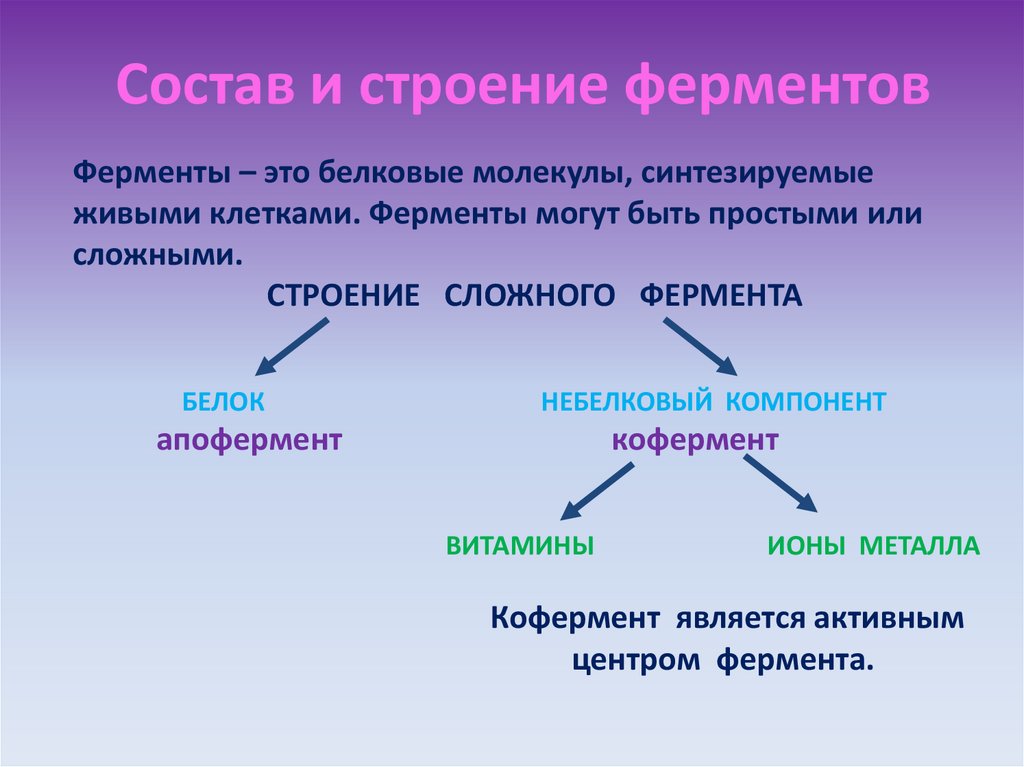 Такая способность возникает в результате формирования промежуточного продукта при связывании антитела с антигеном (имитация переходного комплекса E-X ферментативной реакции).
Такая способность возникает в результате формирования промежуточного продукта при связывании антитела с антигеном (имитация переходного комплекса E-X ферментативной реакции).
Строение ферментов, О.А. Тимин — MedFsh.ru
Общая медицинская теория
ОМТ или Общая медицинская теория — это раздел, в котором размещены теоретические материалы, известные в любом медицинском вузе. Раздел ОМТ в пределах определенного вуза нужен для того, чтобы адаптировать такие материалы под тематические планы вуза. На данный момент тематических планов от пользователей нам не поступало.
К оглавлению: О.А. Тимин «Лекции по общей биохимии»
Давно выяснено, что все ферменты являются белками и обладают всеми свойствами белков. Подобно белкам они делятся на простые и сложные.
Простые ферменты состоят только из аминокислот – например, пепсин, трипсин, лизоцим.
Сложные ферменты (холоферменты) имеют в своем составе белковую часть, состоящую из аминокислот – апофермент, и небелковую часть – кофактор. Кофактор, в свою очередь, может называться коферментом или простетической группой. Примером могут быть сукцинатдегидрогеназа (содержит ФАД), аминотрансферазы (содержат пиридоксальфосфат), пероксидаза (содержит гем).
У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора. Кофактор участвует в связывании субстрата или в его превращении. Для осуществления катализа необходим полноценный комплекс апобелка и кофактора, по отдельности катализ они осуществить не могут.
Аллостерический центр (allos – чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы (называемой активатором или ингибитором, а также эффектором, модулятором, регулятором) вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции. В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество (см «Регуляция активности ферментов»).
В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество (см «Регуляция активности ферментов»).
Аллостерические ферменты являются полимерными белками, их активный и регуляторный центры находятся в разных субъединицах.
Изоферменты
Изоферменты – это молекулярные формы одного и того же фермента, возникшие в результате небольших генетических различий в первичной структуре фермента, но катализирующие одну и ту же реакцию.
Изоферменты отличаются сродством к субстрату, максимальной скорости катализируемой реакции, разной чувствительности к регуляторам (ингибиторы и активаторы), условиям работы (оптимум pH и температуры).
Как правило, изоферменты имеют четвертичную структуру, т.е. состоят из двух или более субъединиц. Например, димерный фермент креатинкиназа представлен тремя изо- ферментными формами, составленными из двух типов субъединиц: M (англ. muscle – мышца) и B (англ. brain – мозг). Креатинкиназа-1 состоит из субъединиц типа B, локализуется в головном мозге, креатинкиназа-2 – по одной М и В субъединице активна в миокарде, креатинкиназа-3 – две М-субъединицы, специфична для скелетной мышцы.
muscle – мышца) и B (англ. brain – мозг). Креатинкиназа-1 состоит из субъединиц типа B, локализуется в головном мозге, креатинкиназа-2 – по одной М и В субъединице активна в миокарде, креатинкиназа-3 – две М-субъединицы, специфична для скелетной мышцы.
Еще одним примером изоферментов является группа гексокиназ, которые присоединяют фосфатную группу к глюкозе и вовлекают глюкозу в реакции клеточного метаболизма. Из четырех изоферментов выделяется гексокиназа IV (глюкокиназа) которая отличается от остальных изоферментов низким сродством к глюкозе и нечувствительностью к ингибированию продуктом реакции.
Также существует пять изоферментов лактатдегидрогеназы (ЛДГ) – фермента, участвующего в обмене глюкозы. Отличия между ними заключаются в разном соотношении субъединиц Н (англ. heart – сердце) и М (англ. muscle – мышца). Лактатдегидрогеназы типов 1 (Н4) и 2 (H3M1) присутствуют в тканях с аэробным обменом (миокард, мозг, корковый слой почек), обладают высоким сродством к молочной кислоте (лактату) и превращают его в пируват.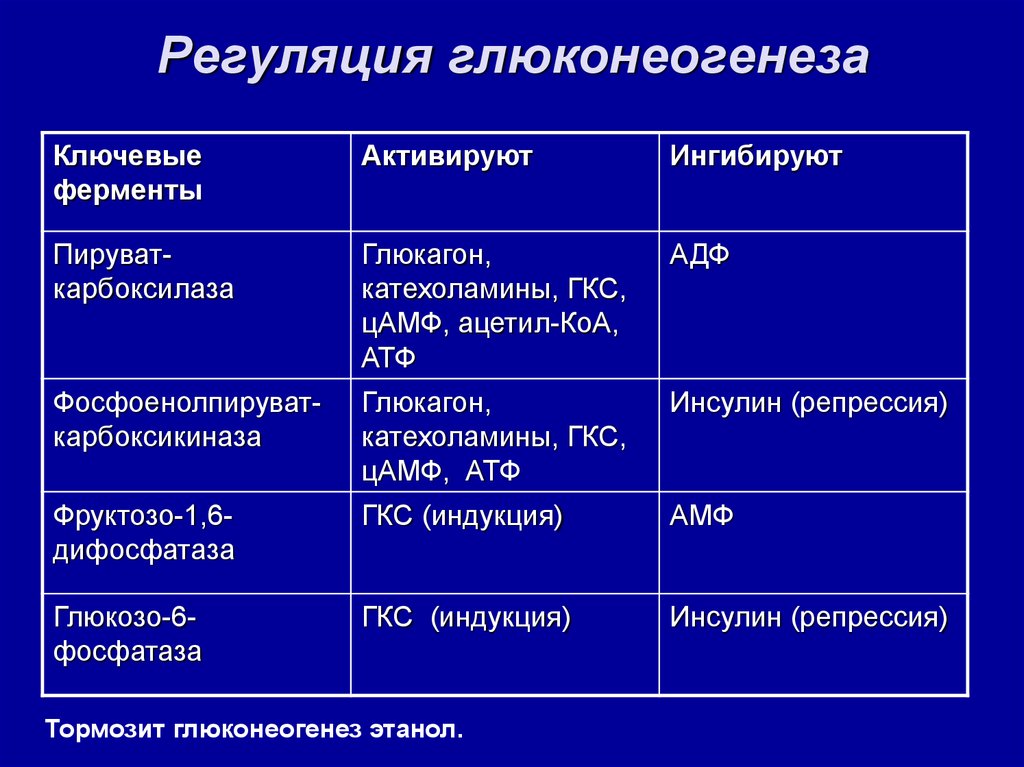 ЛДГ-4 (H1M3) и ЛДГ-5 (М4) находятся в тканях, склонных к анаэробному обмену (печень, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2).
ЛДГ-4 (H1M3) и ЛДГ-5 (М4) находятся в тканях, склонных к анаэробному обмену (печень, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2).
Мультиферментные комплексы
В мультиферментном комплексе несколько ферментов (например, Е1, Е2, Е3) прочно связаны между собой в единый комплекс и осуществляют ряд последовательных реакций, в которых продукт реакции непосредственно передается на следующий фермент и является только его субстратом. Благодаря таким комплексам значительно ускоряется скорость превращения молекул.
К ним относятся, например, пируватдегидрогеназный комплекс, превращающий пируват в ацетил-SКоА, α-кетоглутаратдегидрогеназный комплекс, превращающий α-кетоглутарат в сукцинил-SКоА, комплекс под названием «синтаза жирных кислот» (или пальмитатсинтаза), синтезирующий пальмитиновую кислоту.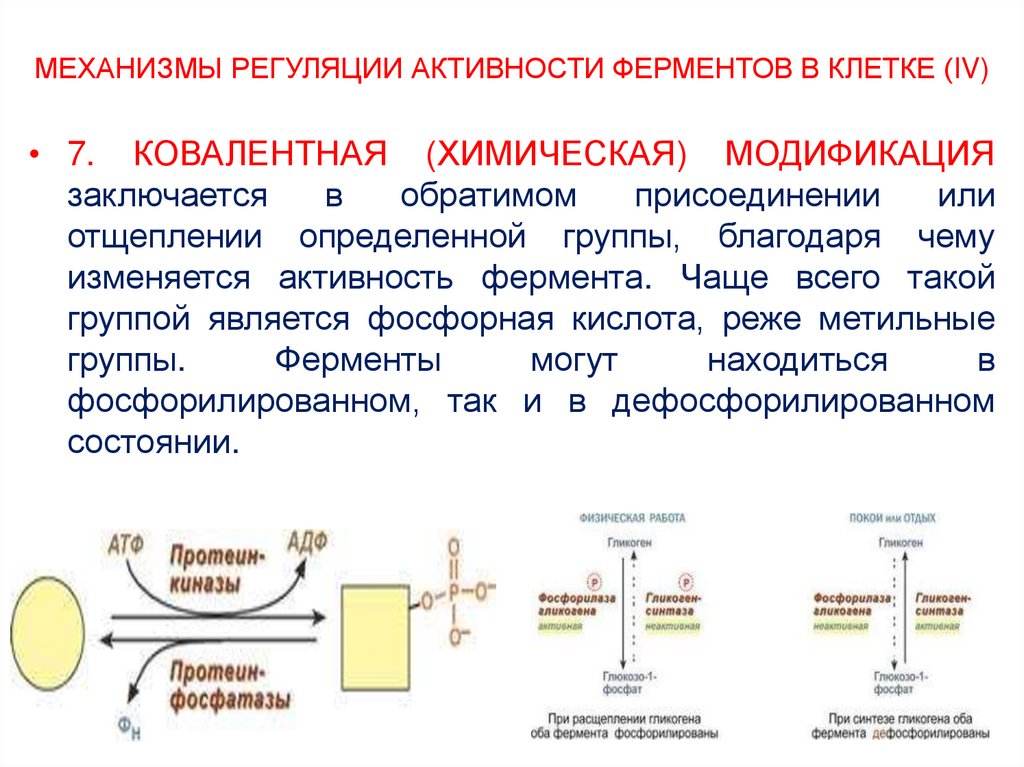
Белки и комплексы электрон-транспортной цепи
Все ресурсы по биохимии
6 Диагностические тесты
289 практических тестов
Вопрос дня
Карточки
Learn by Concept
← Предыдущая 1 2 Следующая →
Биохимия Помощь »
Катаболические пути и метаболизм »
Углеводный обмен »
Электронный транспорт и окислительное фосфорилирование »
Белки и комплексы электрон-транспортной цепи
Какие комплексы цепи переноса электронов будут нарушены при дефиците железа?
Возможные ответы:
Комплекс I и IV
Комплекс II и IV
Комплекс I и II
Комплекс III и IV
Правильный ответ: 90916 5 II0 I0 40 I0
Объяснение:
Комплекс I (NADH-CoQ редуктаза) содержит железо-серные белки, а комплекс II (сукцинат-CoQ редуктаза) содержит как гем, так и железо-серные белки. Таким образом, дефицит железа может поставить под угрозу функцию комплексов I и II. Другие ферментные комплексы не содержат железосодержащих белков, поэтому они не будут нарушены дефицитом железа.
Таким образом, дефицит железа может поставить под угрозу функцию комплексов I и II. Другие ферментные комплексы не содержат железосодержащих белков, поэтому они не будут нарушены дефицитом железа.
Сообщить об ошибке
Какой комплекс электрон-транспортной цепи будет нарушен при дефиците меди?
Возможные ответы:
Комплекс I
Комплекс IV
Комплекс III
Комплекс II
Правильный ответ:
Комплекс IV
Комплекс IV
Объяснение:
Комплекс IV (цитохромоксидаза) содержит два центра меди, и , таким образом, дефицит меди может привести к потере функции ферментного комплекса IV. Другие ферментные комплексы не содержат меди, поэтому дефицит меди не повлияет на их работу.
Сообщить об ошибке
Каков будет самый немедленный результат, если комплекс II электрон-транспортной цепи внезапно перестанет работать?
Возможные ответы:
Увеличение гликолиза
СОКАРЕНИЕ СУКТИКА В митохондриальной матрикске
Гликогеновый распад
Увеличение ионов водорода в митохондриальном пространстве 9000
Правильно Правильно.
Правильный:
. Правильный. в митохондриальном матриксе
Объяснение:
Комплекс II цепи переноса электронов катализирует следующую реакцию:
Он использует фермент сукцинатдегидрогеназу . Непосредственным результатом потери функции этого комплекса будет накопление сукцината, поскольку эта молекула больше не может окисляться до фумарата. Множество проблем, которые могут возникнуть, возникают из-за того, что этот важный этап цикла лимонной кислоты не может двигаться вперед.
Сообщить об ошибке
Какая реакция цикла Кребса осуществляется в электрон-транспортной цепи?
Возможные ответы:
Fumarate to Malate
Сукцинат для фумарата
Цитрат для изоцитрата
Сукцинил-коа для сукцината
Правильный Ответ:
40004
.
Объяснение:
Превращение сукцината в фумарат — единственная реакция, происходящая вне нормального цикла Кребса. Комплекс II цепи переноса электронов имеет фермент, известный как сукцинатдегидрогеназа. Этот фермент отвечает за превращение сукцината в фумарат. Фумарат возвращается в цикл, где затем окисляется до малата, продолжая цикл. Все остальные перечисленные реакции цикла Кребса происходят во внутреннем матриксе митохондрий; тогда как превращение сукцината в фумарат происходит на внутренней митохондриальной мембране.
Сообщить об ошибке
АТФ-синтаза работает посредством __________.
Possible Answers:
an acetyl-CoA gradient across the outer mitochondrial membrane
a proton gradient across the outer mitochondrial membrane
a proton gradient across the inner mitochondrial membrane
an acetyl-CoA gradient across the inner mitochondrial мембрана
Правильный ответ:
протонный градиент через внутреннюю митохондриальную мембрану
Объяснение:
АТФ-синтаза использует протонный градиент через внутреннюю мембрану для образования АТФ. АТФ-синтаза по существу похожа на вращающийся двигатель. Протонный градиент служит инициацией АТФ-синтазы. Когда протоны перемещаются из внешней митохондриальной матрицы обратно в митохондриальную матрицу, они обеспечивают механическую энергию для включения насоса. При включении насоса АТФ-синтаза использует единицу АДФ и неорганического фосфата для образования одной молекулы АТФ. Это делается для каждых трех оборотов АТФ-синтазы.
АТФ-синтаза по существу похожа на вращающийся двигатель. Протонный градиент служит инициацией АТФ-синтазы. Когда протоны перемещаются из внешней митохондриальной матрицы обратно в митохондриальную матрицу, они обеспечивают механическую энергию для включения насоса. При включении насоса АТФ-синтаза использует единицу АДФ и неорганического фосфата для образования одной молекулы АТФ. Это делается для каждых трех оборотов АТФ-синтазы.
Сообщить об ошибке
Комплекс IV электрон-транспортной цепи __________.
Возможные ответы:
несет непосредственную ответственность за производство АТФ из ADP и неорганического фосфата
направляет электрон на кислород с образованием воды
включает в себя сукцинату дегидрогеназу
.
Направляет электрон на кислород с образованием воды
Объяснение:
Комплекс IV также известен как цитохром с-оксидаза, поскольку он принимает электроны от цитохрома с и направляет их на четырехэлектронное восстановление кислорода с образованием двух молекул воды. АТФ-синтаза непосредственно отвечает за образование АТФ, используя одну единицу АДФ и одну единицу неорганического фосфата вместе с протонной движущей силой (PMF). Комплекс II также известен как сукцинатдегидрогеназа, которая отвечает за одну из реакций цикла Кребса: превращение сукцината в фумарат. В результате этой реакции образуется одна молекула . Комплекс I также известен как дегидрогеназа, поскольку он окисляет кофермент.
АТФ-синтаза непосредственно отвечает за образование АТФ, используя одну единицу АДФ и одну единицу неорганического фосфата вместе с протонной движущей силой (PMF). Комплекс II также известен как сукцинатдегидрогеназа, которая отвечает за одну из реакций цикла Кребса: превращение сукцината в фумарат. В результате этой реакции образуется одна молекула . Комплекс I также известен как дегидрогеназа, поскольку он окисляет кофермент.
Сообщить об ошибке
Комплекс I электрон-транспортной цепи __________.
Возможные ответы:
несет непосредственную ответственность за образование воды
. Включает в себя сукцинату дегидрогеназу
. Электроны в кислород
отвечает за принятие электронов из NADH
Правильный ответ:
. Ответственный за Примечание за согласие за согласие. электроны из NADH
Объяснение:
Комплекс I также называют редуктазой NADH-кофермента Q (CoQ), поскольку он переносит 2 электрона от NADH к CoQ. Комплекс I ранее был известен как НАДН-дегидрогеназа. Этот комплекс связывает НАДН и поглощает два электрона. Последним этапом этого комплекса является перенос двух электронов по одному на CoQ. Процесс переноса электронов от NADH к CoQ с помощью комплекса I приводит к общему переносу протонов со стороны матрикса внутренней митохондриальной мембраны в межмембранное пространство, где концентрация ионов водорода увеличивается, создавая протон-движущую силу, которая используется АТФ-синтазой. .
Комплекс I ранее был известен как НАДН-дегидрогеназа. Этот комплекс связывает НАДН и поглощает два электрона. Последним этапом этого комплекса является перенос двух электронов по одному на CoQ. Процесс переноса электронов от NADH к CoQ с помощью комплекса I приводит к общему переносу протонов со стороны матрикса внутренней митохондриальной мембраны в межмембранное пространство, где концентрация ионов водорода увеличивается, создавая протон-движущую силу, которая используется АТФ-синтазой. .
Сообщить об ошибке
Комплекс II электрон-транспортной цепи __________.
Возможные ответы:
. Включает дегидрогеназа
, включающий сукцинату дегидрогеназу
отвечает за принятие электронов от
имеет цитохром С.
Объяснение:
Комплекс II цепи переноса электронов обычно является частью как цепи переноса электронов, так и цикла Кребса. Именно сукцинатдегидрогеназа осуществляет превращение сукцината в фумарат в цикле Кребса.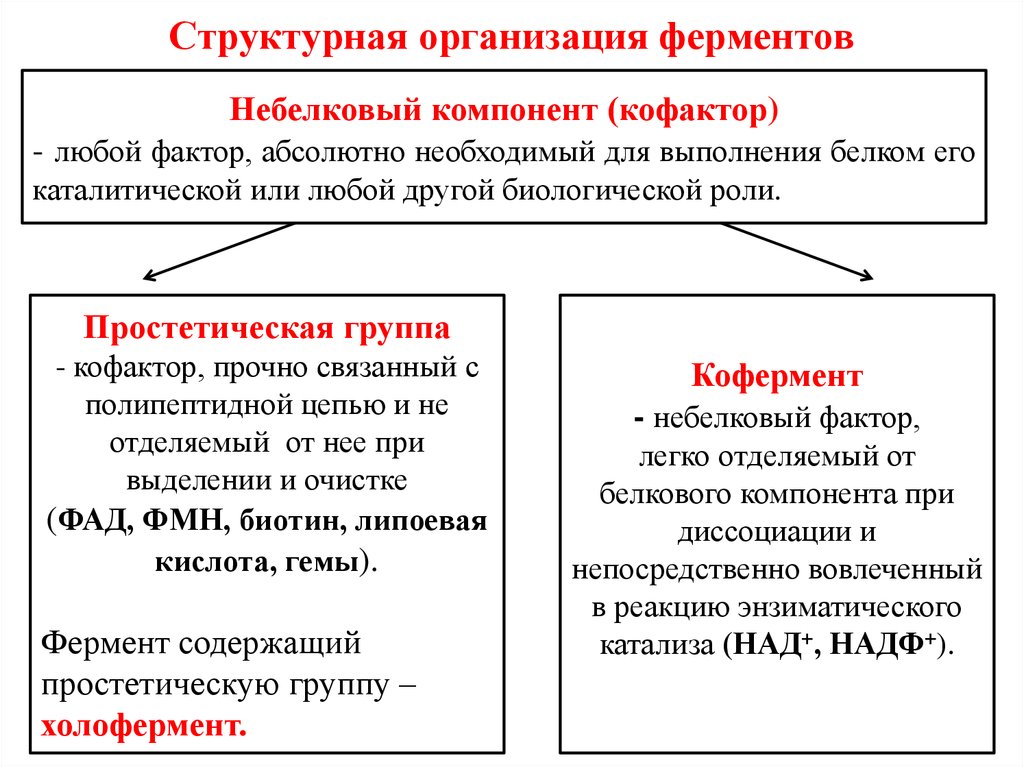 Единственный фермент цикла лимонной кислоты, являющийся интегральным мембранным белком. Превращение сукцината в фумарат приводит к образованию . затем переносит свои электроны по одному через комплекс II. Конечным этапом этого комплекса является перенос двух электронов по одному на кофермент Q.
Единственный фермент цикла лимонной кислоты, являющийся интегральным мембранным белком. Превращение сукцината в фумарат приводит к образованию . затем переносит свои электроны по одному через комплекс II. Конечным этапом этого комплекса является перенос двух электронов по одному на кофермент Q.
Сообщить об ошибке
В комплексе II электрон-транспортной цепи какой кофермент(ы) в основном окисляется?
Возможные ответы:
и только
только
только
Объяснение:
Комплекс II цепи переноса электронов обычно является частью как цепи переноса электронов, так и цикла Кребса. Именно сукцинатдегидрогеназа осуществляет превращение сукцината в фумарат в цикле Кребса. Единственный фермент цикла лимонной кислоты, являющийся интегральным мембранным белком. Превращение сукцината в фумарат приводит к образованию . затем переносит свои электроны по одному через комплекс II. Конечным этапом этого комплекса является перенос двух электронов по одному на кофермент Q.
Конечным этапом этого комплекса является перенос двух электронов по одному на кофермент Q.
Сообщить об ошибке
Какова роль убихинона в цепи переноса электронов?
Возможные ответы:
Убихинон является последней стадией восстановления кислорода до воды.
Убихинон принимает электроны непосредственно от
Убихинон переносит электроны от первого ферментного комплекса ко второму ферментному комплексу
Правильный ответ:
Убихинон переносит электроны от первого ферментного комплекса ко второму ферментному комплексу
Объяснение:
Убихинон выполняет функцию переноса электронов при окислительном фосфорилировании от первого комплекса ферментов ко второму комплексу ферментов. Он не получает электроны ни от ни напрямую.
Сообщить об ошибке
← Предыдущая 1 2 Следующая →
Уведомление об авторских правах
Все ресурсы по биохимии
6 Диагностические тесты
289 практических тестов
Вопрос дня
Карточки
Learn by Concept
Ферментно-субстратный комплекс: определение и примеры
Ферментно-субстратный комплекс Определение
Ферментно-субстратный комплекс представляет собой временную молекулу, образующуюся при идеальном контакте фермента со своим субстратом. Без своего субстрата фермент имеет несколько иную форму. Субстрат вызывает конформационные изменения или изменение формы, когда субстрат входит в активный центр. Активный центр – это участок фермента, способный образовывать слабые связи с субстратом. Это изменение формы может заставить две или более молекулы субстрата объединиться или разделить отдельные молекулы на более мелкие части. Большинство реакций, которые используют клетки, чтобы остаться в живых, требуют, чтобы действия ферментов происходили достаточно быстро, чтобы быть полезными. Эти ферменты непосредственно закодированы в ДНК организма.
Без своего субстрата фермент имеет несколько иную форму. Субстрат вызывает конформационные изменения или изменение формы, когда субстрат входит в активный центр. Активный центр – это участок фермента, способный образовывать слабые связи с субстратом. Это изменение формы может заставить две или более молекулы субстрата объединиться или разделить отдельные молекулы на более мелкие части. Большинство реакций, которые используют клетки, чтобы остаться в живых, требуют, чтобы действия ферментов происходили достаточно быстро, чтобы быть полезными. Эти ферменты непосредственно закодированы в ДНК организма.
Комплекс фермент-субстрат чрезвычайно важен по ряду причин. Во-первых, комплекс фермент-субстрат является временным. Это означает, что после изменения субстрата он больше не может связываться с ферментом. Продукты высвобождаются, и фермент готов к другой молекуле субстрата. Один фермент может работать многократно миллионы раз, а это означает, что в каждой клетке требуется лишь небольшое количество фермента.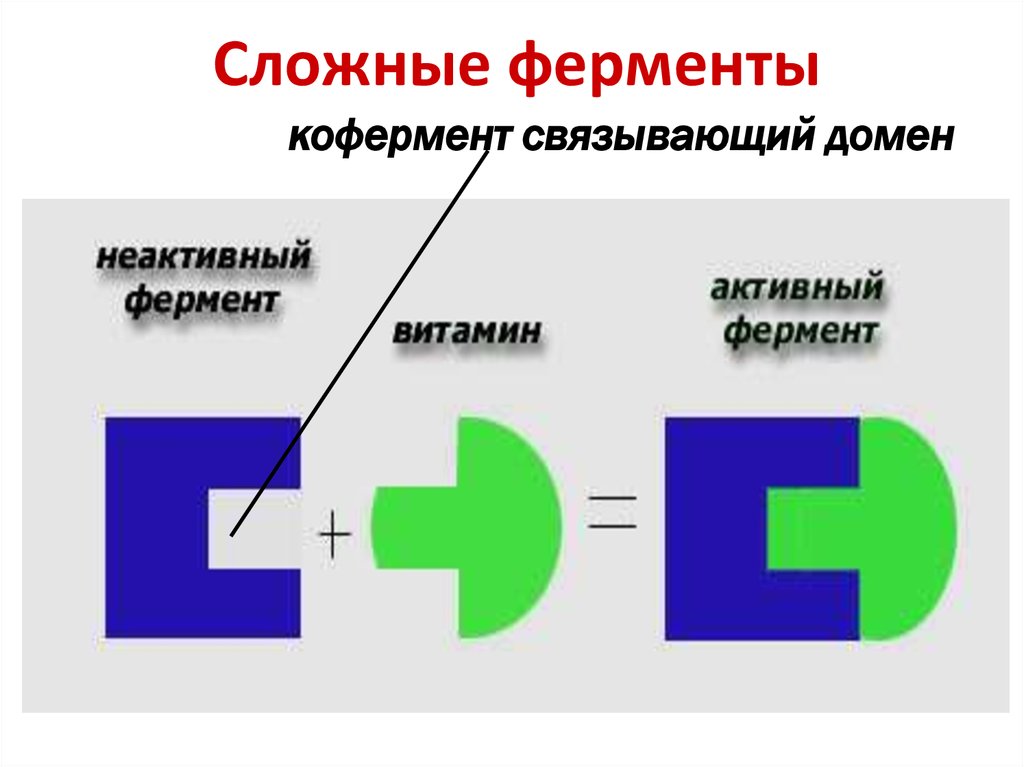
Действие фермента
Ферменты — это сложные молекулы, похожие на маленькие машины, предназначенные для одной цели. Эта длинная цепочка, построенная из цепочки аминокислот, взаимодействует между различными аминокислотами, скручивается и превращается в сложную структуру. Эти структуры могут работать как петли, клинья и всевозможные другие формы, предназначенные для ускорения определенных реакций. Различные мутации приводят к несколько отличающимся формам фермента. При мутациях, полезных для организма, комплекс фермент-субстрат изменяется таким образом, что это влияет на выход продукта или функцию фермента в целом. Это изменение в организме полезно только в том случае, если оно каким-то образом помогает организму воспроизводить больше.
Ферменты обычно называют в честь субстрата, на котором они работают, и имеют суффикс -ase , обозначающий, что они являются ферментами. Каждый фермент имеет определенную специфичность в отношении субстрата, на котором он работает, что определяет, с какими молекулами он может связываться.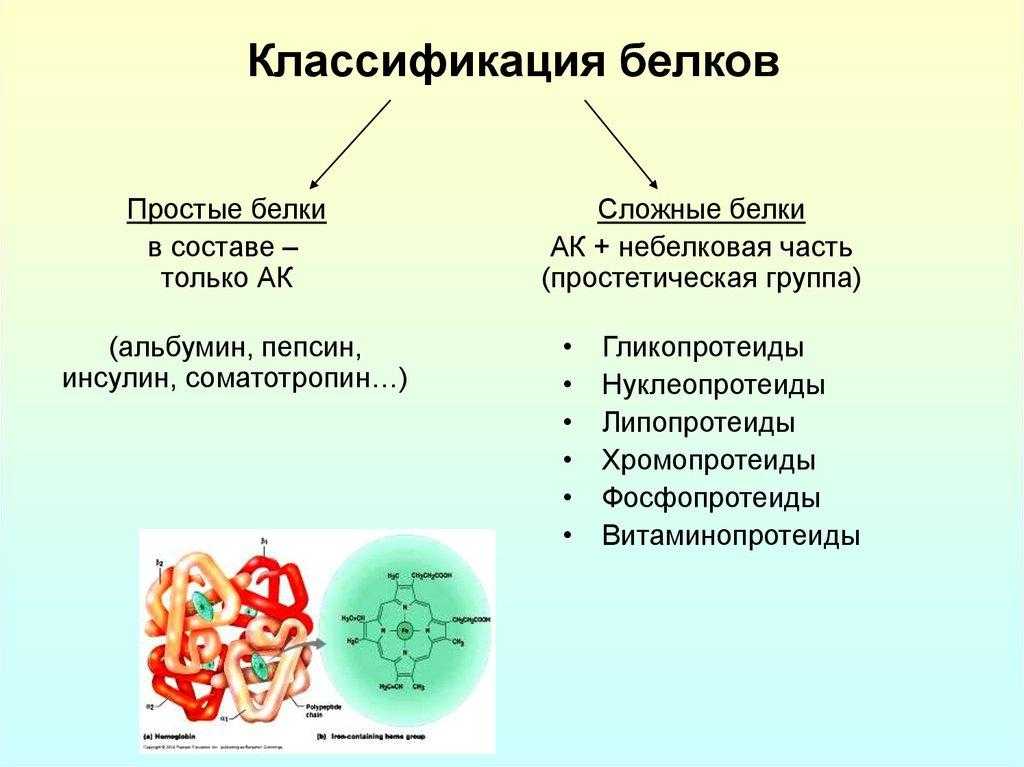 Некоторые молекулы, сходные по структуре с субстратом, могут застрять в активном центре, потому что они не могут вступить в реакцию, предусмотренную ферментом. В этом деформированном фермент-субстратном комплексе конкурентный ингибитор связывается с ферментом и ингибирует его дальнейшее действие. Другие ингибиторы не копируют субстрат, а модифицируют фермент другими способами, так что комплекс фермент-субстрат не может быть образован.
Некоторые молекулы, сходные по структуре с субстратом, могут застрять в активном центре, потому что они не могут вступить в реакцию, предусмотренную ферментом. В этом деформированном фермент-субстратном комплексе конкурентный ингибитор связывается с ферментом и ингибирует его дальнейшее действие. Другие ингибиторы не копируют субстрат, а модифицируют фермент другими способами, так что комплекс фермент-субстрат не может быть образован.
Примеры ферментно-субстратного комплекса
Амилаза и амилоза
Амилоза представляет собой сложный сахар, вырабатываемый растениями. В нашей слюне есть фермент амилаза, используемый для расщепления амилозы. Амилаза использует одну молекулу субстрата амилозы и кофактор одной молекулы воды для получения комплекса фермент-субстрат. Комплекс резко снижает количество энергии, необходимой для запуска реакции, что увеличивает время, в течение которого она происходит. Для распада типичной молекулы сахара потребовались бы миллионы лет, если бы не действие ферментов, таких как амилаза.
На самом деле, ферменты настолько важны для переваривания пищи, которую мы едим, что наше тело вырабатывает фермент почти для каждого вида пищи, который организм эволюционно готов потреблять. Новые продукты плохо перерабатываются, потому что ферменты не успели отрегулировать свою эффективность. Например, современная диета, состоящая из обработанных пищевых продуктов, ведет к эпидемии ожирения, потому что обработанные пищевые продукты богаты легкодоступными питательными веществами, но только тем путям, которые используются для накопления жира. В результате большая часть населения страдает заболеваниями, связанными с лишним весом. Многие диетологи настаивают на более натуральных, цельных, растительных диетах, которые, как правило, поддерживают ферменты, вырабатываемые нашим организмом естественным путем.
Аллостерическая регуляция в ферментах
Хотя быстрое образование комплекса фермент-субстрат важно для большинства реакций, в некоторых случаях важно «выключить» фермент для сохранения энергии или ресурсов.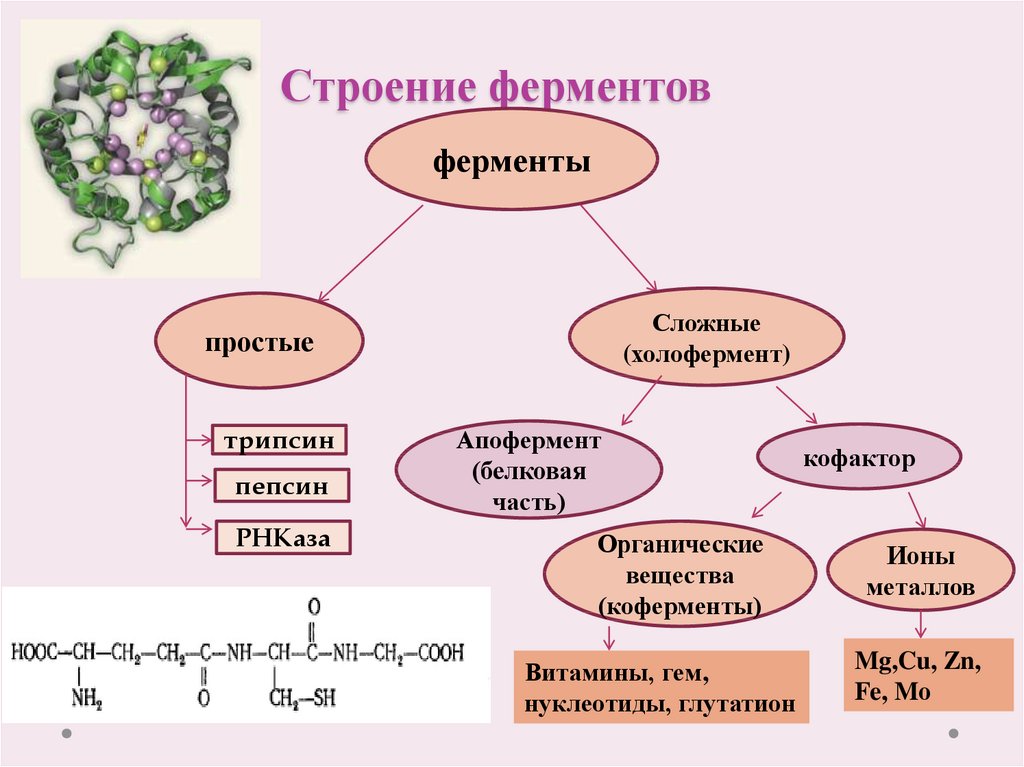 Многие ферменты регулируются таким образом, чтобы обеспечить нужное количество энергии и продуктов. Одним из наиболее важных мест, где это происходит, является производство аденозинтрифосфата (АТФ) или молекулы, обеспечивающей энергией клеточные процессы. Многие ферменты пути, создающего АТФ, инактивируются АТФ. Таким образом, когда вырабатывается слишком много АТФ, фермент отключается. Это известно как торможение по обратной связи , или способность к саморегуляции. Точно так же ферменты могут быть реактивированы в присутствии аденозиндифосфата АДФ, АТФ, который использует фосфатную группу для обеспечения энергией процесса или реакции.
Многие ферменты регулируются таким образом, чтобы обеспечить нужное количество энергии и продуктов. Одним из наиболее важных мест, где это происходит, является производство аденозинтрифосфата (АТФ) или молекулы, обеспечивающей энергией клеточные процессы. Многие ферменты пути, создающего АТФ, инактивируются АТФ. Таким образом, когда вырабатывается слишком много АТФ, фермент отключается. Это известно как торможение по обратной связи , или способность к саморегуляции. Точно так же ферменты могут быть реактивированы в присутствии аденозиндифосфата АДФ, АТФ, который использует фосфатную группу для обеспечения энергией процесса или реакции.
Многие телесные процессы контролируются таким образом, и в этих случаях фермент-субстратный комплекс может быть образован только при наличии соответствующих молекул. Многие из кофакторов, активирующих ферменты, представляют собой витамины, минералы и другие неорганические молекулы, присутствующие в рационе.
- Фермент – Белковые катализаторы, снижающие энергию активации и ускоряющие биологические реакции.

- Субстрат – Молекула или атом, на который действует фермент.
- Энергия активации – Энергия, необходимая для начала реакции.
- Катализатор – Любая молекула или вещество, снижающее энергию активации конкретной реакции.
Тест
1. Угарный газ представляет собой опасную молекулу для вдыхания. В кровотоке угарный газ может проникать в клетки и прочно связываться с железом, кофактором важного фермента в производстве АТФ. Без этой функции фермента АТФ не может быть образован. Почему фермент не работает?
A. Комплекс фермент-субстрат не может быть образован.
B. Угарный газ препятствует попаданию кислорода в легкие.
C. Он все еще может, как только он вызывает реакцию угарного газа.
Ответ на вопрос № 1
Правильно . Без полного формирования комплекса фермент-субстрат с соответствующим субстратом субстрат не может быть трансформирован. Монооксид углерода связывается с кофактором железа, немного изменяя форму фермента и заставляя его больше не принимать свой субстрат, отчасти потому, что молекула монооксида углерода мешает. Фермент не сможет воздействовать на молекулу, но в конечном итоге она будет высвобождена.
Монооксид углерода связывается с кофактором железа, немного изменяя форму фермента и заставляя его больше не принимать свой субстрат, отчасти потому, что молекула монооксида углерода мешает. Фермент не сможет воздействовать на молекулу, но в конечном итоге она будет высвобождена.
2. Некоторые ферменты производят один продукт из двух молекул субстрата. В этих ферментах субстраты загружаются в активный центр, образуется фермент-субстратный комплекс и высвобождается один продукт. Может ли этот процесс происходить в обратном порядке, например, может ли продукт загружаться в фермент и разделяться на два продукта?
A. Да
B. Нет
C. Зависит от фермента
Ответ на вопрос №2
B правильно. Нет, фермент не может работать в обратном направлении. Фермент можно рассматривать как инструмент. Хотя вы можете использовать топор, чтобы расколоть бревно, вы не можете использовать его, чтобы снова сложить бревно.